2. Клеточная мембрана
Клеточная мембрана оформлена в виде трех моно-, би- или полимолекулярных слоев, где периферические слои составлены: белками, а средний - липидами.
В бимолекулярном слое липидов два ряда молекул соединены гидрофобными концами, а гидрофильные концы обращены к белку (Zalokar, 1965).
Фосфолипиды, входящие в состав клеточных мембран, представляют собой фосфоглицериды жирных кислот, содержащие полярные головные группы в форме положительно заряженных холина или этаноламина, отрицательно заряженного серина или; нейтрального гликофосфолипида с гидрофильной головной группой, как в фосфатидилинозите (рис. 7.1). Сочетание в плазматических мембранах фосфоглицеридов с белками может формироваться в виде трех-, четырех-, многослойных образований с фибриллярными и глобулярными белками, полярными и неполярными белковыми структурами или головными группами фосфолипидов (рис. 7.2).
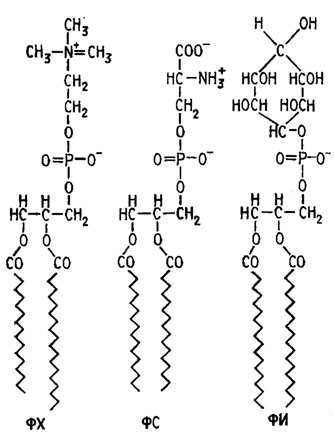
Рис. 7.1. Фосфоглицериды клеточных мембран: ФХ - фосфатидилхолин; ФС - фосфатидилсерин; ФИ - фосфатидилинозит (Либберт, 1976)
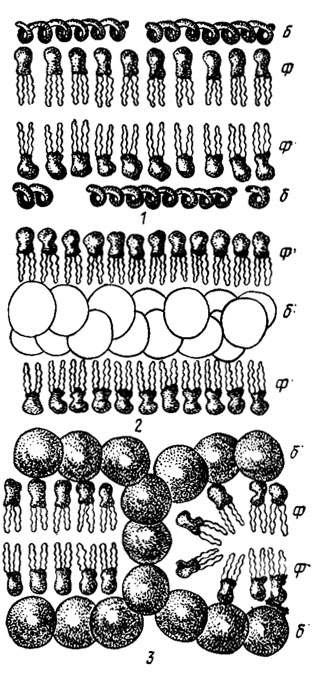
Рис. 7.2. Структуры клеточных мембран, составленных: 1 - из фосфолипидов и фибриллярных белков; 2 - из фосфолипидов и нейтральных глобулярных белков; 3 - из фосфолипидов и полярных глобулярных белков; Ф - фосфолипиды; Б - белки (Либберт, 1976)
Электронная микроскопия и косвенные методы исследования по проницаемости клеток для воды (Solomon, 1961; Rotstein, 1965), антисептиков и антибиотиков (Becker, 1968) по проницаемости мембран для синтетических полисахаридов показали наличие в мембранах многочисленных пор разнообразного размера, характерного для каждой крупной таксономической группы организмов. Сложность строения клеточных мембран грибов была показана на примере их строения у дрожжей, обнаруживаемого с помощью сканирующего электронного микроскопа и техники замораживания и скалывания. В толще мембраны выявляются многочисленные гексагональные структуры и система тяжей, пролиферирующих в область клеточной оболочки (Фрей-Висслинг, 1976; Burnett, 1968).
Основное назначение клеточных мембран, которые в молодых клетках гиф грибов тесно связаны с оболочкой (Беккер, 1956), состоит в регуляции активной проницаемости клеток для поступающих из среды веществ.
Структура клеточных мембран может меняться в зависимости от жирно-кислотного состава ее липидов (Н. В. Усольцева, В. А. Усольцева, 1980). Преобладание в ее составе ненасыщенных жирных кислот придает ей свойства флюидности, а насыщенных - ригидности, что в свою очередь сказывается на интенсивности протекающего в плазмалемме синтеза экзоферментов (Borris, 1981). Доказательством наличия периодических процессов уплотнения и разжижения клеточных мембран у грибов могут служить наблюдения над составом жирных кислот в мицелии Neurospora (Brody, Martins, 1976), у которой количество ненасыщенных жирных кислот колеблется в пределах суточного цикла роста от 37 до 83%.
Функцией липидных слоев, включающих стероиды, является сохранение структуры клеточных мембран и создание барьеров против бесконтрольного поступления и выделения из клеток макромолекул и ионов. Ряд синтезов в процессе обмена грибов может протекать в пределах мембран, например синтез полисахаридных и хитиновых оболочек и экзоферментов (Borris, 1981), а также формирование путем инвагинации мембраны целого ряда клеточных органелл.
|
ПОИСК:
|
При использовании материалов проекта активная ссылка обязательна:
http://gribochek.su/ 'Библиотека о грибах'