Грибы и процесс почвообразования
Почвенные грибы так же, как высшие растения и животные, населяющие почву, принимают активное участие в почвообразовательных процессах, сущность которых заключается в разрушении и создании органического вещества. В этом и состоит основная роль грибов в почве.
Уже отмечалось, что грибы, в том числе и почвенные, являются гетеротрофными организмами, т. е. нуждаются в готовых органических веществах. Они находят их в природе в виде органических остатков или в виде других живых организмов, т. е. существуют или как сапротрофы, или как паразиты растений, животных, других грибов. Иногда невозможно провести резкую грань между этими типами существования, так как они бывают связаны переходами.
Большинство почвенных грибов-сапротрофов отличается широкой амплитудой приспособления к источникам пищи, поэтому один и тот же вид гриба может иногда поселяться на органических остатках различного происхождения. В то же время на одном и том же субстрате, содержащем органические вещества, могут одновременно развиваться различные виды грибов, и тогда между ними иногда складываются очень сложные взаимоотношения взаимопомощи или конкуренции. Чаще бывает так, что процесс разложения органического вещества осуществляется целым комплексом грибов, не говоря уже о других организмах, принимающих участие в этом процессе. Имеются среди почвенных грибов и узкие "специалисты", приспособившиеся к каким-то своеобразным субстратам, где у них нет конкурентов.
В природе ежегодно происходит отмирание различных растений и их частей. Основными разрушителями растительных остатков являются грибы. Это обусловлено в первую очередь кислой реакцией растительных субстратов, которая благоприятствует росту и развитию грибов, и наличием у грибов соответствующих ферментов. Если бы в природе не существовало грибов, вся поверхность Земли вскоре оказалась бы покрытой остатками растений.
В составе растительных остатков по массе преобладают материалы клеточной стенки растений, т. е. клетчатка, или целлюлоза, гемицеллюлозы и лигнин - сложные соединения, в состав которых входит углерод. Следовательно, грибам принадлежит очень существенная роль в круговороте углерода.
Весь процесс разрушения остатков корневых систем растений или их частей происходит непосредственно в почве. Однако в лесу он идет иначе, чем на лугу или в поле, и осуществляется иными грибами. Причина этого заключается в том, что для каждого из перечисленных местообитаний характерны свои комплексы грибов, определяемые различиями физико-химических свойств почвы, влажности, температуры и т. д. Бывает так, что иногда процессу разрушения подвергается живой корень, если на него нападает почвенный гриб-паразит. Осенний опенок, например, является опасным паразитом древесных пород, особенно осины, березы и других деревьев. Он образует хорошо выраженные тяжи из гиф мицелия, протягивает их вдоль корня, и как только такой тяж наткнется на поврежденный участок корня (повреждения часто вызываются животными или насекомыми), гифы тяжа в этом месте тотчас же внедряются в корень, распространяются в нем, а тяж тянется дальше, вновь находит слабое место, и опять гифы входят внутрь корня. Наконец сопротивление дерева ослабляется под действием гиф, и гриб быстро распространяется уже не только в корне, но и в стволовой части, вызывая периферическую белую гниль, которая обнаруживается в стволе иногда на высоте до 3 м, а иногда и выше. Молодые деревья погибают очень быстро после внедрения паразита, а более взрослые могут болеть в течение нескольких лет. Листья их преждевременно желтеют и опадают, а осенью (с конца августа и иногда до заморозков) в основании стволов могут уже появляться плодовые тела опенка. Дерево усыхает, гибнет, ломается от ветра, а на оставшихся частях стволов и пнях развиваются группы плодовых тел гриба. Корни других деревьев могут быстро заразиться грибницей при контакте с корнями уже зараженных. Тяжи, или ризоморфы, опенка могут переходить и на здоровые корни через почву или растительные остатки.

Молодая елочка, погибшая от поражения опенком осенним
Не менее опасный паразит корней деревьев - корневая губка, которая попадает в корни тоже в виде мицелия из почвы; иногда ею заражаются свежесрубленные пни при попадании на них спор этого гриба. Споры прорастают затем в мицелий, внедряющийся в пень, а оттуда он проникает в почву и заражает другие деревья. Для борьбы с этим паразитом наряду с корчеванием зараженных пней был предложен метод биологической борьбы - нанесение на поверхность свежесрубленного пня споровой суспензии какого-либо хорошо и быстро растущего сапротрофного гриба, который распространяется на пне и в его толще, т. е. быстро захватывает субстрат и, таким образом, предотвращает возможность заражения пня корневой губкой.
Нечто подобное происходит в том случае, когда корневая система дерева поражается сначала не такими сильными паразитами, как корневая губка или опенок, а более слабыми, например видами рода фузариум. Обычно вслед за ними в отмирающий корень внедряются грибы-сапротрофы (аспергиллы, пенициллы и др.), быстро растущие и образующие много спор, что позволяет им активно захватывать субстрат. Они усваивают легкодоступные вещества клеток растений, главным образом сахара. Затем другие грибы (ризопус, триходерма) используют более труднодоступные гемицеллюлозы и крахмал. Паразит, не выдерживая конкуренции с этими грибами, уступает им поле боя. В процессе разрушения указанные грибы образуют органические кислоты (лимонную, яблочную и др.), что создает благоприятные условия для развития грибов следующей стадии. Процесс разрушения завершают грибы, воздействующие на еще более прочные материалы клеточной стенки - целлюлозу, или клетчатку, и лигнин. Целлюлозу разрушают виды рода хетомиум из сумчатых грибов, виды родов триходерма, стахиботрис, дикоккум и др. из дейтеромицетов и многие базидиальные грибы.
Лигнин, составляющий от 18 до 30% одревесневших тканей растений, наиболее устойчивый компонент растительных остатков, разрушается главным образом базидиальными грибами, живущими в подстилке, - говорушками, коллибиями, негниючниками, миценами и др. Многие из них разрушают одновременно и целлюлозу и лигнин. Исследованиями установлено, что в процессе разложения растительных остатков (комплексном процессе разрушения лигнина) в различных типах леса участвуют и некоторые дейтеромицеты - фузариум, триходерма, стемфилиум, альтернария.
Существенным компонентом растительных тканей является пектин, который пропитывает и склеивает оболочки клеток стеблей, плодов, корней, семян многих растений. Ферменты, разлагающие пектин, встречаются у различных видов бактерий и у грибов, в том числе почвенных мукоровых, дейтеромицетов (аспергиллов, пенициллов, вертициллов, фузариумов, ботритис, склеротиний и др.). Известная на Руси с давних времен так называемая росяная мочка льна и конопли основана как раз на том, что при расстилании растений на лугах во влажную погоду, при обильной росе, на них происходит массовое развитие микроскопических грибов, в том числе почвенного мукорового гриба - мукора зимнего, ферменты которого, растворяя пектин, разъединяют волокна растений. Пектолитический фермент одного из почвенных грибов рода пеницилл используется, в частности, для осветления фруктовых соков.

Мицены - разрушители лигнина и целлюлозы растительных тканей
Растительные остатки в лесу накапливаются в виде подстилки, биомасса которой в хвойном лесу, например, составляет иногда 300 ц/га, а в широколиственном примерно 150 ц/га. В подстилке лиственного леса различают слои: верхний, состоящий из опада, только слегка тронутого разложением; средний, в котором растительные остатки полуразложились; нижний, сильно разложившийся слой, в котором, помимо органического вещества, можно обнаружить и примесь минеральных частиц. В верхнем слое подстилки присутствует ряд грибов - слабых паразитов, которые попадают сюда вместе с листьями, поселившись на них тогда, когда они были живыми или по крайней мере находились на ветвях дерева. Эти грибы представлены видами родов микосферелла, кладоспориум, альтернария, ауреобазидиум, фома и др. В среднем, наиболее биогенном слое их сменяют другие грибы - подстилочные сапротрофы из базидиальных грибов и почвенные мукоровые грибы (из родов мортиерелла, тиегемелла), сумчатые (виды хетомиум) и дейтеромицеты (главным образом пенициллы). И, наконец, в нижнем слое помимо видов, характерных для предыдущего слоя, присутствует уже свой набор видов (аспергиллы, виды хризоспориум и др.), завершающих процесс разложения. Мы только для примера назвали некоторые виды грибов - участников этих процессов, так как даже в различных типах леса (не говоря уже о специфике процесса разложения растительных остатков в поле и степи) на разных этапах разложения образуются разные комплексы грибов.
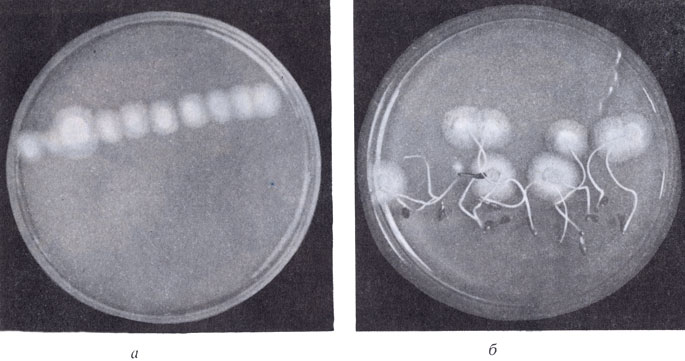
Рост характерного для почв хвойного леса гриба мортиерелла: а - без проростков ели (мицельный рост); б - с проростками ели (колоннии больше и спороносят)
Процессы разложения в подстилке и почве очень сложны, идут в несколько этапов и осуществляются на этих этапах разными видами грибов, т. е. происходит, как говорят, сукцессия видов грибов, набор которых определяется всей экологической обстановкой, начиная с характера субстрата и кончая типом почвы, физико-химическими условиями и т. п. В частности, на основании приведенных выше цифр, характеризующих мощность подстилки хвойного и широколиственного леса, уже можно сделать вывод о том, что в хвойном лесу процесс разложения идет значительно медленнее, что связано с характером опада. Процесс разложения подстилки в значительной степени зависит также от влажности - необходимого условия развития грибов.
Процессы разложения так называемой деловой древесины (шпалы, крепежный лес в шахтах и т. п.) тоже часто происходят в почве. Основные разрушители ее - базидиальные грибы, но, как выяснилось работами последних лет, и здесь немаловажную роль выполняют микроскопические почвенные грибы. Дело в том, что такая древесина в целях защиты от грибов-древоразрушителей обычно пропитывается специальными антисептиками, а микроскопические почвенные грибы, поселяясь на ней, разрушают эти . антисептики, после чего открывается доступ в нее основным древоразрушителям.
Интересны связи и взаимоотношения между компонентами в сукцессиях грибов. Уже отмечалось, что быстрорастущие и появляющиеся на первых этапах разложения древесных материалов грибы образуют органические кислоты, чем создают более благоприятные условия для развития грибов последующих стадий разложения.
Некоторые виды микроскопических грибов могут использовать для своей жизнедеятельности вещества, образующиеся после отмирания мицелия грибов предшествующей стадии, в частности разрушать вещества клеточных стенок грибов. Это виды родов мукор, мортиерелла, аспергиллы и др.
Определенные взаимоотношения складываются между грибами и почвенными бактериями. Известно, что базидиальные грибы - подстилочные сапротрофы благоприятствуют развитию аммонифицирующих бактерий, что же касается микроскопических почвенных грибов, то они проявляют антагонизм к целлюлозоразру-шающим бактериям, являющимся их конкурентами в этом процессе. На почве конкуренции между организмами у грибов выработались в процессе эволюции такие средства подавления своих противников, как антибиотики. Вместе с тем специально проведенные опыты показали, что микроскопические почвенные грибы обычно не угнетают развитие грибов - подстилочных сапротрофов.
Процессы разложения органического вещества происходят не только в почве или на ее поверхности, но и в наземных условиях и примерно по такой же, как и описанная выше, схеме.
Почвенные микроскопические грибы принимают участие в круговороте не только углерода, но и азота. Остатки растений и животных, содержащие азот, попадают в почву, где разрушаются бактериями, актиномицетами и грибами. Например, в черноземах из 10 т соединений азота, приходящихся на 1 га, 90% азота находится в форме органических соединений- белков, мочевины мочевой кислоты и др. Особенно активные участники превращений этих соединений - мукоровые грибы, виды аспергиллов, триходермы и др. Почти все они способны также усваивать азот аммонийных соединений, но далеко не все - нитратный азот. Это соединение очень хорошо усваивается почвенным грибом аспергиллом черным.
Известно, что при недостатке в почве безазотистого органического питания (углеводов) в ней накапливается значительное количество аммиака, образующегося в результате деятельности мукоровых грибов при разложении аммонийных соединений азота. Если в почве углеводов много, мукоровые грибы могут энергично усваивать растворимые минеральные соединения азота (нитраты, те же аммонийные соединения) и переводить их в белки своего тела. Благодаря этому уменьшается вымывание азотистых веществ из почвы, так как они закрепляются в мицелии грибов.
В почву попадают и такие специфические вещества, как перья птиц, копыта и рога животных, остатки их волосяного покрова, хитиновые оболочки насекомых и других животных. На них развиваются специфические комплексы грибов, в конечном счете приводящие их к полной деградации. Птичьи перья, рога, копыта, волосы состоят из кератина, который под воздействием грибов размягчается и растворяется. Соответствующие грибы можно "выманить" из почвы, если на ней поместить отмытые и обезжиренные волосы, а потом уже можно выделить эти грибы на питательной среде с целью их изучения. При просмотре таких волос под микроскопом при сильном увеличении в них можно видеть гифы грибов. Из грибов, разлагающих кератин, можно назвать онигену лошадиную, развивающуюся на копытах и рогах, онигену воронью - на перьях и волосах, а также типичного почвенного разрушителя кератина - кератиномицеса айеллои. Чаще всего такие грибы встречаются в почвах богатых органическими веществами - в местах выпаса скота, садах.
Не менее прочное, чем кератин, и трудноразлагаемое вещество - хитин. Он содержится в покровных тканях насекомых и многих других обитателей почвы, в клеточных стенках мицелия грибов. Основные разрушители хитина в почве - актиномицеты или, как их раньше называли, лучистые грибки (теперь их относят к бактериям). Некоторые микроскопические почвенные грибы тоже обладают этой способностью, и их можно также выделять, пользуясь методом приманок. Относятся эти грибы к видам родов мукор, мортиерелла, аспергилл, хетомиум, тиелавиопсис и др. Особенно активная роль в разрушении хитина приписывается грибу аспергиллу дымчатому.
Есть гриб, который приурочен к почвам, пропитанным нефтью, и часто развивается на керосине, креозоте, дизельных топливах, различных смазках. Это кладоспориум смолистый, или керосиновый гриб.
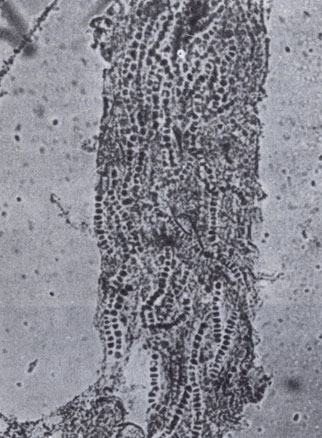
Почвенный гриб кератиномицес айеллои внутри волоса
Почвенные грибы участвуют в процессах почвообразования, не только разрушая различные вещества, попадающие в почву. Грибы способствуют и оструктуриванию почвы или, иначе говоря, образованию деятельного или активного перегноя. Особенно важна роль грибов, живущих в тяжелых глинистых и суглинистых почвах, так как оструктуривание их грибами увеличивает аэрируемость и улучшает снабжение их водой.
В конце XIX в. образование комочков почвы приписывалось исключительно деятельности дождевых червей, позже на первый план выступили физико-химические свойства почвы, т. е. образование почвенной структуры связывалось с действием растворимых солей кальция, магния, калия, обладающих осаждающей способностью, а затем снова возникли биологические теории структурообразования, в том числе было доказано участие в этом процессе грибов. Оказалось, что выделяемые почвенными грибами вещества как бы склеивают почвенные частички друг с другом. Этому же способствуют и сами нити грибницы, оплетающие и пронизывающие частички. Даже тогда, когда грибница отмирает, продукты ее разложения продолжают выполнять эту функцию.
Многие почвенные грибы являются хорошими структурообразователями, но, как показали опыты, особенно сильной агрегирующей способностью обладают некоторые мукоровые (виды родов мукор, ризопус, абсидия), сумчатые (виды рода хетомиум), некоторые дейтеромицеты (виды родов триходерма, аспергилл, фузариум). Часто внесение этих грибов в почву вместе со свободно живущей азотфиксирующей почвенной бактерией азотобактером хроококкум способствовало более хорошему росту бактерии, что также улучшало структуру почвы.
Деятельность грибов в почве не ограничивается, однако, теми процессами, о которых было сказано. Существует мнение, что образование и разрушение гумуса также связано с деятельностью почвенных грибов и в процессе их жизни и после гибели мицелия в окружающую среду обильно выделяются вещества, которые могут принимать участие в синтезе гумуса. В частности, некоторые аспергиллы и пенициллы, темноокрашенные дейтеромицеты выделяют пигменты и антибиотические вещества, способные соединяться с другими азотсодержащими продуктами разложения органических веществ, в том числе белками (аминокислотами, пептидами), продуктами деградации лигнина, и образовывать комплексы, сходные с гуминовыми кислотами, входящими в состав гумуса. Особое значение в этом процессе имеют как раз темные пигменты, присущие многим почвенным грибам, особенно дейтеромицетам. Оказалось, что эти пигменты (меланины) по ряду химических и физических свойств очень сходны с гуминовыми кислотами.
Некоторые почвенные грибы способны и разлагать гумус, использовать его в качестве единственного источника углерода и азота. Например, присутствующий в некоторых почвах пеницилл частый освобождает в ходе этого процесса азот, который может быть использован высшими растениями.
В состав гумуса входит фосфор - элемент, необходимый для нормального роста и развития высших растений. Грибы, в частности пенициллы, образующие органические кислоты, активно участвуют в превращении нерастворимого фосфора в растворимый и, следовательно, доступный высшим растениям.
Таким образом, почвенные грибы представляют собой чрезвычайно активную группу, принимающую важное участие в процессах почвообразования. Своими ферментами они содействуют разложению различных материалов, попадающих в почву, и переводят часть переработанных веществ в компоненты своего тела. Другие части или отлагаются в почве, меняя ее химический состав и свойства, или используются и перерабатываются другими организмами. Многие почвенные грибы воздействуют и на минеральную основу почвы органическими кислотами, которые образуются и накапливаются при разложении этими грибами различных составных частей растительных клеток. Особенно активно накопление этих кислот протекает в подзолистых лесных почвах, в которых грибы представлены многочисленными видами пенициллов. В результате этого отдельные зольные элементы переходят в растворенное состояние, происходит разложение минералов, образование комплексных соединений, перемещающихся в нижележащие слои почвы, - развивается процесс подзолообразования.
Нельзя не упомянуть и о роли почвенных грибов в токсикозе почв. Некоторые виды грибов выделяют в почву ядовитые вещества - токсины, и почва при этом часто становится токсичной для высших растений. Частый случай токсикоза почв - почвоутомление. Оно заключается в том, что растения на таких почвах угнетаются, снижают урожайность. Особенно часто это бывает, когда в течение многих лет на одном и том же месте возделывается какая-нибудь одна культура. Токсикоз почв отражается и на почвенных микроорганизмах, причем иногда это совпадает с угнетением развития высших растений. Как выяснилось, наибольшее количество токсичных видов среди пенициллов. Токсичны и некоторые виды родов триходерма, фузариум, альтернария, аспергиллы и др. Все они выделяют вещества, могущие подавлять прорастание семян растений.
Длительность сохранения токсинов в почве различна. В опытах одни из них быстро или вымывались, или инактивировались, другие сохранились в течение нескольких дней.
Наибольшее количество токсических видов почвенных грибов содержат подзолистые, серо-бурые лесные и бурые таежные почвы.
Некоторые почвенные грибы вырабатывают токсины, действующие на животных и на людей.
Известно, что некоторые виды почвенных грибов образуют вещества, стимулирующие рост растений. Одни из этих веществ - гиббереллины - образует гриб из рода фузариум. Он представляет собой конидиальную стадию сумчатого гриба - гиббереллы, откуда и получили свое название вещества. Впервые они были обнаружены японским исследователем Е. Куросавой в связи с изучением болезней риса, среди которых наиболее загадочной считалась "болезнь дурных побегов", или бакане, распространенная в Японии, Китае, Индии и ряде других стран. (Междоузлия риса, пораженного этой болезнью, сильно вытягиваются, а все растение очень сильно ветвится.) Причиной заболевания риса оказался фузариум, а затем были выделены и вещества, вызывавшие симптомы заболевания. Впоследствии выяснилось, что гиббереллины способны стимулировать у различных растений не только рост побегов, листьев, в меньшей степени корней, но и ускорять цветение и плодоношение. Вещества эти, а их известно уже несколько, адсорбируются почвой и могут сохраняться в нестерильной почвенной среде физиологически активными в течение более 1,5 мес.
В последние годы гиббереллины используют, в частности, в тех случаях, когда требуется увеличить у технических культур длину стеблей, а следовательно, и волокон, ускорить цветение или выгонку семенников у декоративных растений.
Часто наблюдается задержка или угнетение прорастания спор грибов, находящихся в почве. Такое явление называется фунгистазисом и наиболее отчетливо выражено в почвах с большим количеством микроорганизмов и недостатком питательных веществ. Объясняется оно тем, что в таких почвах особенно обостряется конкуренция между обитателями почвы и активный мицелий грибов может разрушаться другими микроорганизмами. Споры в этом отношении более стойки и могут сохраняться какое-то время в состоянии покоя до тех пор, пока условия не станут более благоприятными для их прорастания. Следовательно, состояние фунгистазиса полезно для грибов. Еще лучше грибы могут переживать неблагоприятный период в почве в виде таких структур, как хламидоспоры, зиготы, склероции и т. п. Наличие таких структур особенно характерно для грибов-паразитов растений (видов из родов фузариум, вертициллиум, биполярис, склеротиния, аскохита), у которых такие структуры могут сохраняться некоторое время в почве при отсутствии восприимчивых к ним высших растений-хозяев. Таким образом, фунгистазис представляет собой вынужденный покой, возникший и закрепившийся в процессе эволюции как полезный для сохранения вида. Если искусственно прервать этот покой и вызвать прорастание покоящихся структур или спор, например до появления на поле восприимчивых растений или до развития тех стадий, на которых они восприимчивы, то таким приемом можно понизить вероятность поражения растения соответствующим паразитом. Эта же цель может быть достигнута и обратным путем, т. е. длительной задержкой прорастания покоящихся структур до тех пор, пока высшее растение-хозяин пройдет в своем развитии ту стадию, на которой оно особенно подвержено заражению паразитом, и таким образом избегнет заражения.
|
ПОИСК:
|
При использовании материалов проекта активная ссылка обязательна:
http://gribochek.su/ 'Библиотека о грибах'