Глава 3. Источники углерода в питании грибов и углеродный обмен
1. Источники углерода в питании грибов
Гетеротрофность грибов выражается в основном в их потребности в органических источниках углерода, по отношению к которым они практически всеядны и не используют только некоторые пластики и "неразрушаемые" детергенты (Perlman, 1965). Отсюда вытекает их негативное практическое значение как разрушителей всевозможных строительных и других материалов, пищепродуктов, изделий и моторных топлив.
Однако при этом выявляются и полезные свойства таких грибов, заключающиеся в возможности их использования для переработки разнообразных отходов промышленности и всевозможных, органических остатков, накапливающихся в биосфере.
Используемый грибами круг источников углерода очень велик. Неполный список органических соединений, используемых грибами: как единственный источник углерода, включает 155 соединений и состоит из 56 углеводов, сахарных кислот и спиртов производных Сахаров, 43 аминокислот и других азотсодержащих соединений, 40 органических кислот и их сложных эфиров и 16 стероидных соединений и алкалоидов.
Обычно алифатические соединения более питательны для грибов, чем ароматические. Исключение для Aspergillus niger составляет хинная кислота, но у нее другое распределение двойных связей в молекуле, при котором разрыв кольца протекает значительна легче, чем у ароматических соединений типа бензола.
Питательная ценность углеводов обычно учитывается по газообмену и по массе образующегося на среде с ними мицелия. Особенно хорошо усваиваются из числа сахаров находящиеся на первом месте по питательной ценности гексозы, содержащие шесть атомов углерода, в первую очередь глюкоза и фруктоза. Фруктоза усваивается несколько слабее, чем глюкоза, и еще слабее галактоза.
Однако такие водные грибы, как например Leptomitus, не используют глюкозу, фруктозу, галактозу и сахарозу. Также некоторые штаммы головневого гриба Ustilago striiformis при пересеве их со среды с сахарозой на среду с глюкозой три-четыре недели приспосабливаются к глюкозе, пока не приобретают способность усваивать ее достаточно интенсивно. Из дисахаридов сахароза потребляется лучше, чем лактоза. Пентозы менее доступны для грибов, чем гексозы, и среди них ксилоза доступнее арабинозы. Некоторые виды рода Fusarium (например, F. oxysporum, вызывающий вилт табака) хорошо усваивают ксилозу, а также мальтозу, но экономический коэффициент потребления Сахаров у него выше на левулозе (Wolf, 1955). Для Fusarium moniliforme сахара распределяются в порядке следующего ряда интенсивности их использования: глюкоза > фруктоза > l-арабиноза.
На втором месте по питательности после моносахаров находятся ди-, три- и полисахариды. Однако есть исключения, например возбудитель фузариозного вилта хлопчатника F. oxysporum f. vasinfectum лучше развивается на средах с белками и крахмалом, чем на среде с глюкозой. При росте его на средах с целлюлозой, липидами и ДНК наблюдается длительная лаг-фаза, которая сокращается только при добавлении в среду глюкозы. Также ряд оомицетов, например некоторые сапролегниевые грибы (Apodachlya brachynema) и ряд видов Phytophthora, предпочитают моносахарам крахмал и декстрин. Allomyces использует только мальтозу, сахарозу и декстрин (Лилли, Барнеп, 1953). Многие сапролегниевые грибы вообще не потребляют углеводы, например Leptomitus lacteus, для которого источником углерода служат жирные кислоты и аминокислоты.
Правило предпочтения гексоз действительно для большей части почвенных и паразитных грибов. Например, возбудители антракноза виды Colletotrichum и Gloeosporium (Tandom, Agarwala, 1956) хорошо растут на среде с глюкозой, фруктозой, маннозой и рамнозой и особенно интенсивно - на среде с двумя первыми сахарами. Ксилозу и арабинозу хорошо используют не все виды этих родов грибов.
Спирты, производные моносахаров, например маннит, усваиваются хорошо, так же как и сахарные кислоты: глюконовая, глюкуроновая и сахарная. Исключение составляют некоторые дерматофиты, не потребляющие маннита, - признак, которым пользуются в систематике рода Sporotrichum.
На смесях углеводов рост грибов нередко заметно усиливается, что имеет экологическое значение, поскольку в природных условиях им приходится встречаться с гетерогенными смесями источников углерода. Примером усиления роста на смеси Сахаров может служить Aspergillus nigery показывающий массу сухого мицелия на среде с 18 г галактозы - 42,4 мг, на среде с 2 г глюкозы - 145,6 мг, а на среде со смесью из 18 г галактозы и 2 г глюкозы - 577 мг. При сложении эффекта влияния на массу мицелия двух Сахаров должна была бы получиться масса сухого мицелия, равная 188 мг (Steinberg, 1939). Сходные явления наблюдались у видов рода Gloeosporium, которые при росте на смесях из маннита, раффинозы, галактозы и сорбита показывали гораздо более интенсивную споруляцию, чем на средах с каждым из этих веществ в отдельности (Tandom, Agarwala, 1956). На основе улучшающего рост эффекта смесей углеводов производилась композиция целого ряда питательных сред. Например, среда Билай для видов рода Fusarium составлена с включением по 1% глюкозы, сахарозы и крахмала.
Для сред со смесями углеводов и органических кислот установлена определенная очередность потребления их компонентов. Например, Penicillium chrysogenum сначала усваивает в такой среде ацетат, вторыми - лактат и часть глюкозы и в последнюю очередь - лактозу.
Выращивание на среде со смесью углеводов может в ряде случаев способствовать адаптации гриба к определенному сахару, который не может быть использован с помощью конститутивных, т. е. присутствующих в первоначально образовавшемся мицелии энзимов. Примером может служить выращивание грибов на среде с сорбозой, слабо или совсем не поддерживающей рост многих грибов (Lilly, Barnett, 1953).
При выращивании на средах с сорбозой в комбинации с мальтозой или глюкозой, поддерживающими рост грибов из адаптирующейся к сорбозе группы (Choanephora, Mucor, Phycornyces), эти грибы растут и вырабатывают ферменты, нужные для усвоения сорбозы. При этом настолько усиливается рост культур на средах с добавкой сорбозы, что это нельзя объяснить только суммированием питательной ценности обоих Сахаров, как в случае видов Aspergillus, Fusarium и Penicillium.
Неспособная адаптироваться к сорбозе группа грибов (Alternaria, Rhizoctonia и Sclerotinia) оказывается угнетенной в росте на средах со смесью Сахаров, содержащей сорбозу, так как сорбоза способствует торможению их метаболизма.
Питательное значение алифатических кислот возрастает с увеличением длины углеродной цепи до шести углеродов, а именно в форме ряда с увеличивающейся интенсивностью их усвоения:
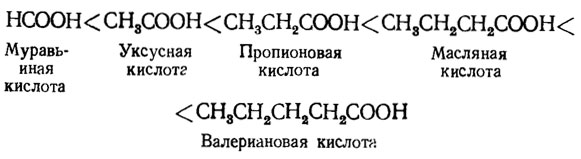
Двухосновные кислоты более питательны, чем одноосновные, а оксикислоты усваиваются лучше, чем обычные, например:

Большую питательную ценность для грибов имеют также и другие дикарбоновые и трикарбоиовые кислоты, содержащие гидроксильные группы, например:
СООНСН(ОН)СН(ОН)СООН
Винная кислота
и СООНС(ОН)СН2СООН
|
СООН
Лимонная кислота
Особенно хорошо усваиваются грибами соединения, содержащие группировки -СН(ОН)-, чем и объясняется высокая питательная ценность оксикислот и сахаров.
Однако запас энергии в окисленных соединениях обычно меньше, чем в менее окисленных. Бедна ею карбоксильная группа -СООН-, что видно из слабого использования таких кислот, как муравьиная НСООН и щавелевая СООН-СООН, которая практически совсем не потребляется и обладает токсичностью.
Для усвоения углеводов и кислот большое значение имеет их пространственная изометрия, что было обнаружено еще Л. Пастером. Исследовавшийся им штамм из рода Penicillium лучше использовал (+) винную кислоту, чем (-) винную, на основе чего оказалось возможным отделить левовращающую винную кислоту из ее рацемической формы.
Некоторые виды грибов потребляют органические кислоты очень слабо, например Pleospora indica, паразит пальмы Cycas revoluta. Она растет только на среде с сахарозой, плохо - на среде с крахмалом и инулином и из двенадцати испытанных органических кислот не использует ни одной (Mandahar, 1965).
Наивысшей питательностью для грибов обладают спирты с шестью атомами углерода (например, маннит); меньшей - трехатомные спирты, как глицерин, и очень слабой - одноатомные типа этилового и метилового. Альдегиды и кетоны используются грибами хуже спиртов.
Жиры (триглицериды жирных кислот) хорошо усваиваются целым рядом грибов, предварительно расщепляющих их на жирную кислоту и глицерин липазами. Из продуктов расщепления жиров грибы раньше всего потребляют глицерин, а потом уже жирные кислоты: пальмитиновую, стеариновую и олеиновую. Жирно-кислотные компоненты жиров очень хорошо используют сапролегниевые грибы (например, Leptomitus lacteus), что связано с их экологией, поскольку они расщепляют в основном субстраты животного происхождения, а также Penicillium chrysogenum и ряд других видов рода Penicillium.
Близок по строению к жирам воск - сложный эфир мирицилового спирта и пальмитиновой кислоты. Использовать его как источник углерода может Aspergillus niger, часто являющийся разрушителем пчелиных сотов. Представители семейства Aspergillaсеае могут также питаться парафином, но лучше они усваивают углеводы.
Хорошо растут на среде с парафиновыми углеводородами некоторые дрожжи, которые служат для получения кормового белка, применяемого в животноводстве. Способность использовать для питания углеводороды свойственна и ряду гифообразующих почвенных грибов: мукоровым из родов Absidia, Mortierella, Cunninghamella, Mucor, видам родов Fusarium, Aspergillus, Penicillium, Spicaria и Trichoderma, которые усваивают углеводороды нефтепродуктов (Билай, Коваль, 1980).
Грибы, расщепляющие углеводороды, разрушают горючее к смазки, являются причиной закупорки масло- и бензопроводов, что может вызывать воздушные аварии, коррозию деталей самолетов и других машин. Из карбюраторов самолетов была выделена культура Cladosporium resinae, устойчивая к углеводородам с высоким октановым числом и способная развиваться на них как на единственном источнике углерода (Nicot, Zakartchenko, 1966). Участие углеводородов в обмене грибов подтверждается обнаружением их среди продуктов метаболизма. Так, например, из хламидоспор Ustilago maydis, U. nuda и Sphacelotheca reiliana было выделено соответственно 42, 56 и 146 видов алканов, представленных нормальными парафинами с длиной цепи от 14 до 37 атомов углерода. Подобные же углеводороды выделены из спор-видов Puccinia, хламидоспор Tilletia и спор целого ряда других грибов (Weete, 1980).
Исследование углеродного питания можно использовать для целей таксономии. Так, Курсанов упоминал о Saccharomyces globosus, не содержащем инвертазы и не использующем сахарозу, который употреблялся для изготовления сладких вин. Недоброд сахара при применении этого вида составлял 20-25%, а содержание спирта в вине порядка 6-7%.
В отношении углеводного питания различаются между собой не только виды грибов, но также их расы и штаммы. Выделенные из различных сортов хлопчатника штаммы возбудителя вилта Fusarium oxysporum f. vasinfectum отличались друг от друга не только по морфологическим признакам, но и по способности использовать различные сахара. Штамм F3 использовал лучше фруктозу, тогда как штамм F1 - мальтозу. Таким образом, физиологические критерии следует использовать в таксономии для изучения филогенетических связей очень осторожно, поскольку огромное значение для формирования сходных потребностей в питании имеет сходная экология. Например, как было установлено, из четырех дереворазрушающих грибов (аскомицетов Xylaria digitata и Нуроxylon rubiginosum, базидиомицетов Polyporus cinnabarinus к P. conchifer) no питанию источниками азота и углерода P. cinnabarinus оказался ближе к Н: rubiginosumt чем к P. conchifer (Rajagopalan, 1965).
Углеводы играют роль не только в процессах роста грибов, на и как стимуляторы прорастания спор и как индукторы биосинтеза адаптивных ферментов. У базидиомицета Schizophyllum commune сахарные спирты активируют прорастание спор в течение экспозиции 30-60 мин, а большая часть Сахаров и гликоген в течение 15-20 ч. Лаг-фаза для прорастания спор этого гриба в присутствии лактозы, сорбозы, раффинозы, трегалозы и мелибиозы составляла от 30 ч до 7 дней, а в присутствии галактозы, инозита, инулина, рамнозы, фукозы и органических кислот базидиоспоры не прорастали даже после 7 дней инкубации (Niederpruem, Dannen, 1966). В качестве индукторов адаптации ферментативной активности в отношении целлюлитических энзимов у 70 исследованных дереворазрушающих грибов могут служить целлобиоза и лактоза, особенно активные в присутствии дрожжевого экстракта, а также некоторые белки (Johnson, 1966). Ароматические соединения, даже содержащие 6 атомов углерода, усваиваются очень слабо.
Из соединений углерода наиболее простого строения грибы могут усваивать углекислоту. Однако единственным источником углерода для гетеротрофных по типу их питания грибов СО2, видимо, служить не может. Она усваивается только в процессе определенного этапа углеродного обмена при переходе от гликолитического цикла к циклам ди- и трикарбоновых кислот или жирных кислот, включаясь в фосфоэнолпируват при образовании оксалацетата или в ацетальдегид при образовании малоновой кислоты.
Усвоение СО2 грибами было впервые показано с помощью изотопного метода Фостером (1941) на моделях Rhizopus nigricans и Aspergillus niger. Из усвоенной грибом при росте на среде с 5% глюкозы в замкнутом сосуде 14СО2 более 1/3 ее оказалось в составе мицелия, а в анаэробных условиях 25% в образованной км фумаровой кислоте (Лилли, Барнетт, 1953; Фостер, 1950). Показана также способность усваивать углекислоту у Blastocladiella pringsheimii и у дереворазрушающего гриба Coniophora cerebella. Усвоение углерода из СО2 воздуха меланинсодержащими грибами рода Cladosporium рассмотрено в работе Н. Н. Ждановой и др. (1985).
|
ПОИСК:
|
При использовании материалов проекта активная ссылка обязательна:
http://gribochek.su/ 'Библиотека о грибах'