3. Полифосфаты и ДНК в клетках грибов
Из встречающихся в больших количествах неорганических соединений в клеточных оболочках грибов, в частности в оболочках конидий Neurospora, найдено много полифосфатов, связывающих около 11% катионов оболочки и образующих в ней солеподобные соединения хитозанфосфаты. У дрожжей фосфаты связаны с глюкопротеидами, и, кроме того, в их оболочках содержится немного нуклеиновых кислот.
Однако роль полифосфатов как высокоспециализированных макроэргов, организующих специфику энергоснабжения и питания грибов, не сводится только к участию в структуре клеточной оболочки и требует специального рассмотрения.
Прежде всего необходимо заметить, что полифосфаты грибов имеют прямое отношение к гранулам так называемого волютина, образующимся в их вакуолях при действии слабых оснований (например, сильно разбавленного аммиака) или основных красителей.
Волютин грибов имеет непостоянный состав в их онтогенезе, способен растворяться не только в кислотах, но и в слабо подщелоченных солях (например, в 0,1 М NH4NO3, подщелоченном до рН 8,0 и в 2-2,5 М растворе аммиака) (Беккер, 1956). Его накопление согласуется с нарастанием солерастворимой фракции полифосфатов в процессе активного синтеза нуклеиновых кислот (Кулаев, 1975).
Использование цитохимических методов, таких как сравнительная фотосъемка в световом и ультрафиолетовом микроскопе, обработка сформированных волютиновых гранул рибонуклеазой, окраска их пиронином и толуидиновой синей после извлечения из них нейтрального красного этанолом (рис. 1.3) позволили уточнить их состав. В процессе онтогенеза продуцента пенициллина он менялся от состава чисто полифосфатной природы через промежуточные двуслойные структуры к чисто рибонуклеиновому (Дмитриева, Беккер, 1962). Согласно исследованиям Л. Н. Кац (1964), состав предсуществующего волютина изученного ею актиномицета менялся в диаметрально противоположном направлении - от РНК-волютина к полифосфатному. По всем этим данным можно с достаточными основаниями утверждать, что волютин грибов несхож с истинным предсуществующим волютином прокариотов, а скорее схож с криномом животных.
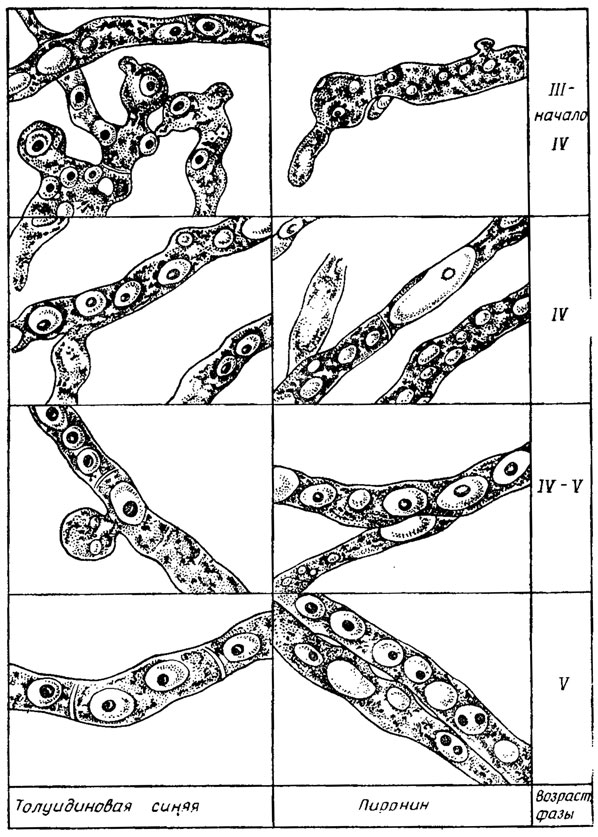
Рис. 1.3. Волютин, сформировавшийся в виде гранул после витальной окраски мицелия Penicillium chrysogenum, из погруженной культуры в его различных возрастных фазах. Обесцвеченные этанолом гранулы в III и в начале IV возрастных фазах окрашиваются в красноватый цвет толуидинблау и не окрашиваются пиронином, т. е. содержат полифосфаты. Начиная с перехода мицелия в V возрастную фазу они окрашиваются пиронином и в синий цвет толуидинбла, т. е. содержат РНК (Дмитриева, Беккер, 1962)
Что полифосфатный обмен грибов отличается от такового у актиномицетов и бактерий, можно видеть также и по синтезирующим и расщепляющим полифосфаты ферментам (Кулаев, 1975). Как видно из прилагаемой схемы (табл. 1.6), у грибов, бактерий и актиномицетов есть общие энзимы этого обмена, такие как полифосфат-фосфогидролаза (ПФГ), полифосфаткиназа (ПФК) и дифосфоглицерат-полифосфаттрансфераза (ДФГ-ПФТ).
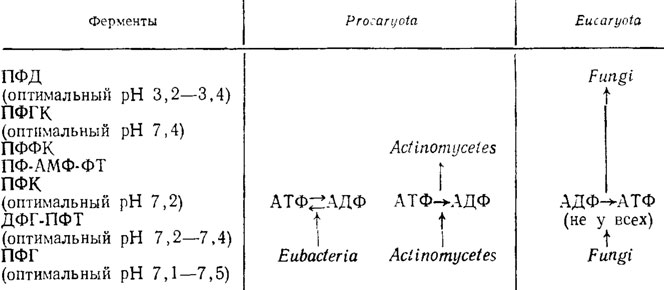
Таблица 1.6. Включение в метаболизм полифосфатов различных ферментов у бактерий, актиномицетов и грибов (Кулаев, 1975) (стрелками показано наличие ферментов в данной группе организмов)
Однако уже такой энзим, как ПФК, переносящий фосфат с АТФ на полифосфат и обратно с полифосфата на АДФ, действует в обоих направлениях только у Eubacteria, а у актиномицетов и грибов только в одном и взаимопротивоположном. У грибов синтез полифосфатов за счет АТФ вообще не возможен.
У форм, причисляемых Н. А. Красильниковым (1970) к актиномицетам, имеются кроме этих еще три энзима: полифосфатглюкокиназа (ПФГК) и полифосфатфруктокиназа (ПФФК), а также полифосфат-АМФ-фосфотрансфераза (ПФ-АМФ-ФТ), отсутствующие у Eubacteria и у грибов. Напротив, грибы обладают, как и в случае многих других их ферментных систем, системой энзимов с действием эндотипа, расщепляющей полифосфаты на разной длины олигомеры в середине их цепи. Эти энзимы действуют в противоположность предшествующим только в кислой зоне рН (оптимальный рН 3,2-3,4) и носят название полифосфатполифосфогидролаз, или полифосфатдеполимераз (ПФД), Они, видимо, имеют адаптивный характер.
Полифосфаты представляют собой чисто неорганические соединения, состоящие из мономеров ортофосфорной кислоты, чаще всего построенные в форме линейных неразветвленных цепей, содержащих от трех, четырех и до 300 мономеров фосфата (Кулаев, 1975): 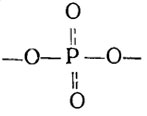
Иногда они, видимо, встречаются в форме двухфрагментиого пирофосфата или циклических метафосфатоз (например, три-, тетра- и гексамерных). Возможны также нестойкие сетчатые структуры и более стойкие сочетания фосфорной кислоты с двух- и трехвалентными металлами, образующими сетчатые структуры с фрагментами ортофосфата, как, например, обнаруженные у грибов (Кулаев, 1975), имеющие функции детоксикаторов избытка свободного ортофосфата и запасников фосфора и металлов (рис. 1.4).
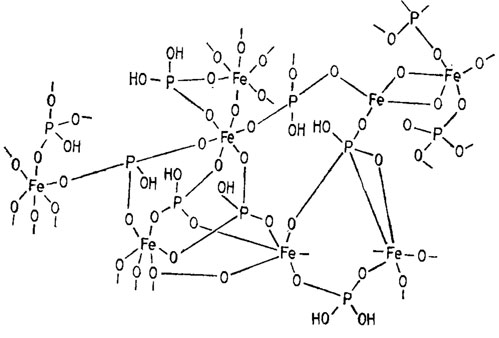
Рис. 1.4. Предположительная структура полимерного ортофосфата железа (Кулаев, 1975)
Наиболее распространенная у грибов и энергетически доступная форма высокомолекулярных полифосфатов представляет собой линейные цепи с очень различным числом фрагментов фосфата, существующие в виде непрерывного ряда соединений от пирофосфата (два фрагмента фосфата) до содержащих 300 фрагментов и более высокополимерных форм. Разделение их на фракции с помощью экстракции разными растворителями является довольно условным, однако дает все же возможность связать эти фракции с определенной их локализацией и функциями в клетке. Обычно насчитывают четыре-пять таких фракций (табл. 1.7).

Таблица 1.7. Характеристика степени полимерности разных фракций полифосфатов пивных дрожжей (Кулаев, 1975)
Согласно способу их экстракции эти фракции носят названия кислоторастворимой (ПФ1, солерастворимой (ПФ2), щелочерастворимых (ПФ3 и ПФ4) и растворимой в хлорной кислоте при нагревании (ПФ5). Средняя длина цепи и средняя молекулярная масса каждой фракции, как можно видеть из результатов анализа фракций из дрожжей (см. табл. 1.7), последовательно возрастают, достигая длины цепи порядка 260n и молекулярной массы около 30000, При этом четыре из перечисленных фракций были обнаружены у всех подробно изученных в этом отношении грибов (табл. 1.8).

Таблица 1.8. Содержание полифосфатов в клетках Endomyces magnusii. Nearospora crassa и в плодовых телах Gyromitra esculenta в мг фосфора на г сухого мицелия (Кулаев, 1975)
В общем итоге полифосфаты были найдены в мицелии грибов всех обследованных классов, а именно у двух миксомицетов, двух видов мукоровых грибов, восьми сумчатых, двух базидиальных, 13 видов несовершенных и 14 видов дрожжей, т. е. у всех из обследованного 41 вида грибов (Кулаев, 1975). По-видимому, наиболее обильно у грибов представлена фракция ПФ2, а в наименьшей степени фракции ПФ3 и ПФ5, однако их относительное количество сильно зависит от вида гриба, условий его роста и возраста культуры.
Биосинтез конденсированных полифосфатов по всем данным начинается в основном с наиболее высокополимерной фракции, а остальные получаются путем ее деградации. Однако возможны и восходящие синтезы за счет пирофосфата, освобождающегося в метаболических процессах, например при синтезе нуклеиновых кислот. Этим процессом, видимо, объясняется синхронное с синтезом РНК у Neurospora crassa накопление фракции ПФ2, сочетающееся с уменьшением содержания наиболее лабильной кислоторастворимой фракции ПФ1 (рис. 1.5).
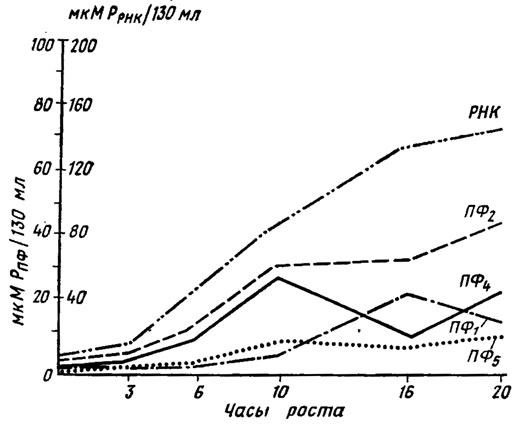
Рис. 1.5. Изменение содержания в мицелии различных фракций полифосфатов и РНК в процессе роста культуры Neurospora crassa в микромолях РО4-3 на 130 мл среды (Кулаев, 1975)
Общие функции конденсированных линейных полифосфатов в клетках грибов весьма схожи с функциями АТФ и состоят в снабжении для метаболических реакций фосфором и энергией, высвобождающейся при разрыве макроэргической связи -О-Р-. Однако у них есть и сильные отличия от АТФ, так как, во-первых, способность их к высокой степени полимеризации дает возможность резервировать большие запасы энергии и фосфора без нарушения гомеостаза клетки (рН, ионной силы, осмотической концентрации и т. д.). Это очень важно для характерных обилием этих соединений низших эукариот (грибов, водорослей и простейших) и прокариот (бактерий, цианей и актиномицетов), так сильно зависимых от, окружающей их среды. Во-вторых, высокомолекулярные полифосфаты обладают определенной специализацией в метаболических процессах. По всей вероятности, наименее специализирована лабильная фракция ПФ1, участвующая как донор энергии во многих ферментативных процессах и синтезах. Фракция ПФ2 специфично связана с синтезом нуклеиновых кислот. Более высокомолекулярные фракции участвуют в транспорте через клеточную мембрану Сахаров, переносчика которых они фосфорилируют, а их синтез связан с синтезом полисахаридов клеточных оболочек. Последнее можно видеть по высоким коэффициентам корреляции между накоплением этих полисахаридов и соответствующих им фракций полифосфатов (табл. 1.9).

Таблица 1.9. Коэффициенты корреляции (r) между скоростями накопления различных фракций полифосфатов и полисахаридов у Saccharomyces carlsbergiensis (Кулаев, 1975)
Проведенные Кулаевым, его учениками и рядом других исследователей работы по изучению локализации различных фракций полифосфатов в клетках грибов позволили обнаружить согласованность между процессами, сопровождающими их биосинтез, и местами их накопления в клеточных органеллах. В протопластах Neurospora crassa, освобожденных ферментом улитки от клеточной оболочки, оказались гидролизованными до оставшегося в протопласте ортофосфата обе наиболее высокомолекулярные фракции полифосфатов (ПФ4 и ПФ5), что указывало на их локализацию на поверхности клеточной мембраны вблизи от оболочки клетки (табл. 1.10).

Таблица 1.10. Содержание высокомолекулярных полифосфатов в клетках, протопластах, ядрах и митохондриях Neurospora crassa. Данные в мг фосфора/г сухого вещества (Кулаев, 1975)
Дальнейшее фракционирование нативных клеточных органелл с помощью ультрацентрифугирования в градиенте сахарозы и их анализ показали наличие в ядре только одной фракции (ПФ2), связанной с биосинтезом РНК. Кислоторастворимая фракция (ПФ1) и большая часть солерастворимой фракции (ПФ2), скорее всего входящие в состав волютина грибов, остались в протопласте, где, как известно, протекает гриколиз; в месте локализации дыхательной цепи, в митохондриях, полифосфатов не оказалось совсем.
Относительно фосфорных соединений необходимо добавить, что, хотя мы и не имеем сейчас точных критериев для суждения о составе нуклеиновых кислот, можно все же утверждать, что они отличаются у грибов от других организмов по их количественной характеристике. Так, по нашим данным (Беккер, Дмитриева и др., 1972), доля ДНК от общего количества фосфорных соединений в мицелии Aspergillus fumigatus не превышает 7-9%, тогда как. доля РНК достигает 65-70% от их количества. Если сравнить обогащенность клетки ДНК у разных организмов, то выясняется, что по этому критерию, так же как и по числу пар нуклеотидов на клетку, грибы находятся на относительно невысоком уровне сложности организации их генома, приближаясь в этом отношении к низшим беспозвоночным, как моллюски или губки. Они далеко отстают от высших растений, сложность организации генома которых сопоставима с таковой у птиц, рыб и ракообразных (Ленинджер, 1974). Однако при сравнении с бактериями сложность генома грибов оказывается выше на целый порядок (грибы - 0,02-0,17; бактерии - 0,002-0,06 мкг ДНК на клетку).
|
ПОИСК:
|
При использовании материалов проекта активная ссылка обязательна:
http://gribochek.su/ 'Библиотека о грибах'