Хищные Гифомицеты (И. И. Сидорова)
Хищничество, редкое среди растений, известно у некоторых грибов. Грибы, способные поймать, убить и использовать в пищу микроскопических животных - нематод, коловраток, простейших или мелких насекомых (Collembola), объединяются в экологическую группу хищных грибов. Большинство представителей этой группы - гифомицеты, однако сюда относятся и представители других таксономических групп - зигомицеты (порядок зоопаговые), грибы рода зоофагус из порядка пероноспоровых класса оомицетов, некоторые хитридиомицеты.
У хищных грибов мицелий развивается в почве, на растительных остатках и других субстратах, но часть питания они получают из тканей пойманной ими жертвы. Тело жертвы представляет для них, как и для хищных животных, только пищу, а не среду обитания, как для паразитов. Захват жертвы хищником (в данном случае грибом) представляет единичный акт, а не процесс совместного существования, как при паразитизме.
Первые описанные в литературе хищные грибы - артроботрис пышный (Arthrobotrys superba) и артроботрис малоспоровый (A. oligospora) - долгое время считали обычными сапрофитными грибами. М. С. Воронин наблюдал образование на мицелии второго из них колец и сетей, описанных им подробно в 1869 г. Позднее, в 1871 г., такие же кольца и сети наблюдал Н. В. Сорокин. Роль этих сетей в улавливании нематод была показана работами немецкого ученого В. Цопфа в конце XIX в.
Однако интенсивное изучение этой своеобразной группы грибов было начато значительно позднее, после появления в 30-х годах нашего века серии работ американского ученого Ч. Дрекслера по морфологии и систематике хищных грибов. За прошедшие с тех пор четыре десятилетия были выяснены многие вопросы таксономии, биологии и распространения хищных грибов.
Вегетативный мицелий хищных грибов состоит из обильно ветвящихся септированных гиф толщиной не более 5-8 мкм. В старых гифах нередко образуются хламидоспоры. На мицелии развиваются различные ловчие приспособления, описываемые ниже. Конидии у хищных грибов развиваются на вертикально стоящих конидиеносцах различного строения и имеют одну или несколько перегородок. У грибов рода артроботрис (Arthrobotrys) конидии двухклетные. Первая конидия образуется бластогенно на вершине конидиеносца, затем ниже ее возникает новая точка роста и развивается новая конидия. Этот процесс повторяется многократно, в результате чего образуется гроздь конидий на вершине конидиеносца, часто утолщенной и бородавчатой. Если в одной из последовательных точек роста происходит пролиферация конидиеносца и этот процесс повторяется, на конидиеносце образуется серия утолщенных узлов, несущих конидии (рис. 246). У представителей рода дактилярия (Dactylaria) на вершине конидиеносца образуется группа многоклеточных конидий, расположенных на стеригмах или бородавчатых выростах конидиеносца, а для видов рода монакроспориум (Monacrosporium) характерно образование на вершине конидиеносца одиночной многоклеточной конидии, часто с более крупной центральной клеткой (рис. 246). Кроме того, к хищным гифомицетам относятся представители родов тридентария (Tridentaria) и трипоспорина (Triposporina) со звездчатыми спорами (рис. 246) и другие грибы.
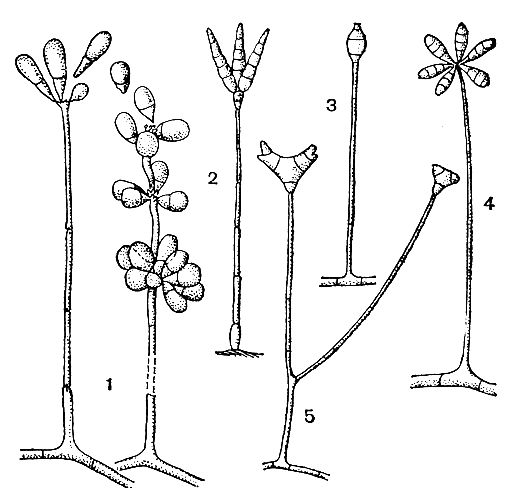
Рис. 246. Конидиальные спороношения хищных грибов: 1 - артроботрис (Arthrobotrys); 2 - тридентария (Tridentaria); 3 - монакроспориум (Monacrosporium); 4 - дактилярия (Dactylaria); 5 - трипоспорина (Triposporina)
Жертвы хищных грибов - обычно сапрозойные нематоды или свободноживущие личинки нематод, патогенных для растений, животных и человека. Реже грибы ловят амеб или других мелких корненожек, а некоторые - мелких насекомых. Таков, например, артроботрис насекомоядный (A. entomophaga), улавливающий представителей коллембола (Collembola).
Часто хищные грибы улавливают животных, значительно превосходящих их по размерам. Размеры нематод (круглых червей), улавливаемых грибами,- 0,1-1,0 мм, а толщина гиф этих грибов не более 8 мкм. Улавливание таких крупных, подвижных и сильных жертв, как нематоды, стало возможно в результате приобретения грибами в процессе эволюции различных специализированных ловчих приспособлений. Строение аппаратов-ловушек у хищных гифомицетов разнообразно, по механизму действия они могут быть трех типов.
Наиболее распространены у хищных гифомицетов клейкие ловушки. В простейшем случае это недифференцированные боковые выросты гиф, покрытые клейким веществом (Arthrobotrys perpasta, Monacrosporium cionopagum). Другие хищные гифомицеты образуют ловчие аппараты в виде маленьких овальных или шаровидных клейких головок, сидящих на ко-ротких двухклетных веточках мицелия (Monacrosporium ellipsosporum, A. entomophaga, рис. 247). Но самый распространенный тип клейких ловушек - клейкие сети, состоящие из большого числа колец. Такие сети образуются в результате обильного ветвления гиф, веточки которых загибаются и анастомозируют с соседними веточками или родительской гифой, образуя сложную трехмерную сеть из многочисленных колец (рис. 247). Поверхность гиф сети покрыта клейким веществом неизвестной природы. Предполагают, что по происхождению оно близко к смолам и гутте, а его биосинтез тесно связан с терпеновым обменом (З. Э. Беккер). Такого типа ловушки известны у многих видов из рода артроботрис, например у часто встречающегося во всех районах земного шара артроботриса малоспорового. Сети часто достигают крупных размеров, и вероятность попадания в них многочисленных и подвижных нематод очень велика.
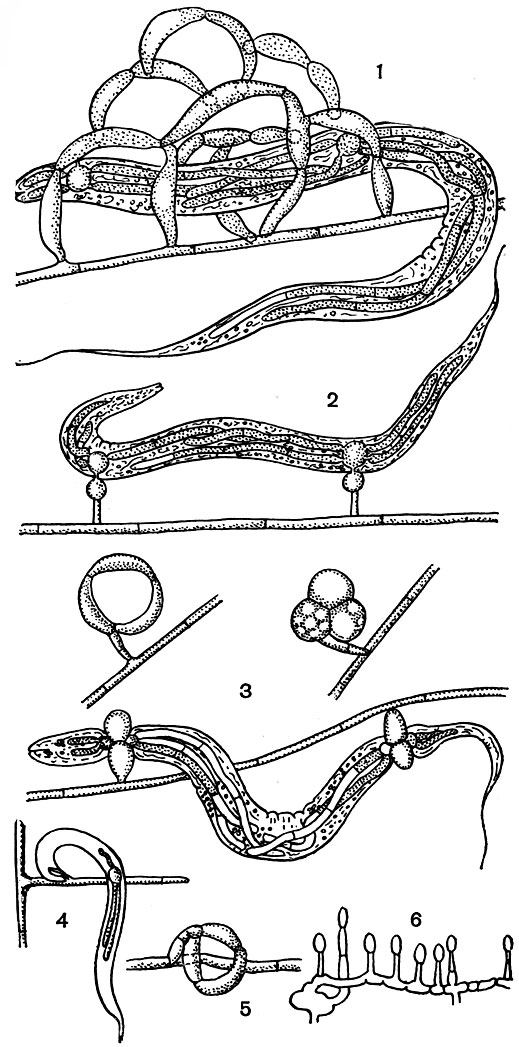
Рис. 247. Типы ловушек хищных грибов: 1 - клейкие трехмерные сети; 2, б - клейкие головки; 3 - сжимающиеся кольца; 4 - клейкие выросты гиф; 5 - несжимающиеся кольца
Процесс улавливания нематоды клейкими сетями напоминает ловлю мух на липкую бумагу. Прикоснувшись к сети, нематода прилипает к ней. Пытаясь освободиться, она активно двигается, извивается и все больше прилипает к сети. Через некоторое время ее движения становятся вялыми, наконец нема-тода перестает двигаться. Вскоре после захвата нематоды клейкой сетью из этой сети развивается гифа, растворяющая кутикулу нематоды и проникающая в ее тело. Часто после прободения кутикулы в теле нематоды образуется так называемая инфекционная луковица, из которой развиваются трофические гифы. Постепенно они заполняют все тело нематоды, и ее ткани теряют свою структуру. Процесс поглощения грибом содержимого тела нематоды продолжается немногим более суток. После этого остается только кутикула нематоды, заполненная трофическими гифами гриба. Крупные, сильные нематоды иногда разрывают ловчие сети и уходят из них, унося на кутикуле прилипшие обрывки гиф. Но и в этом случае нематоды быстро гибнут в результате развития из этих обрывков колец гиф, проникающих в их тело.
У некоторых хищных грибов образуются ловушки в виде колец, лишенных клейкого вещества и действующих механически. Обычно эти кольца состоят из трех изогнутых клеток и расположены на коротких веточках мицелия. В простейшем случае такие ловушки действуют пассивно. Нематода, случайно попав в такую ловушку, пытается пройти сквозь кольцо и застревает в нем. Ловушки этого типа образует дактилярия белоснежная (Dactylaria Candida).
Наиболее интересный тип ловушек хищных грибов - сжимающиеся кольца (рис. 247,3). Внешне они очень похожи на кольца предыдущего типа, однако их внутренняя поверхность тактиосенситивна, т. е. чувствительна к прикосновению к ней (кутикулы нематоды, иглы микроманипулятора). В ответ на раздражение клетки кольца в течение 0,1 сек резко увеличиваются в объеме (примерно в 3 раза) и приобретают шаровидную форму, почти полностью закрывая просвет кольца. Вздутие клеток кольца при обычных условиях необратимо. Если нематода попадает в такую ловушку, она активно захватывается грибом. Проникновение гриба в тело нематоды и питание ее тканями происходят как и в случае с клейкими ловушками. Гибель нематоды в сжимающемся кольце может происходить в результате ее механического сдавливания, так как диаметр ее тела в месте захвата грибом уменьшается вдвое. Сжимающиеся кольца образуются у представителей родов дактилярия, монакроспориум и немногих грибов рода артроботрис.
Механизм действия сжимающихся колец-ловушек до сих пор не изучен полностью. По сведениям Дж. P. Лоутон, замыкание колец хищных грибов стимулируется ацетилхолином. На основании этого она предполагает, что механизм смыкания колец-ловушек сходен с механизмом сокращения мышечного волокна.
Хищные гифомицеты легко выделяются в чистую культуру и растут на питательных средах, но в отсутствие нематод они обычно не образуют аппаратов-ловушек. Если в культуру добавить нематод, то уже через 24 ч в ней образуются ловушки. Стимулировать их образование можно также, добавляя к культуре стерильную воду, в которой жили нематоды. Таким образом, присутствие жертвы или продуктов ее обмена индуцирует у хищника образование ловушек.
Иногда наблюдают неспецифическую индукцию развития ловушек экстрактами из тканей животных, сывороткой крови, ионами СО3 и другими воздействиями. В культуре некоторых нематод были обнаружены вещества, стимулирующие образование ловушек у хищных гифомицетов и получившие название немин. Предполагают, что это низкомолекулярный пептид или аминокислота. Из тела аскарид был получен белок с неминовой активностью. У некоторых хищных гифомицетов, например у артроботриса дактилоидного (A. dactyloides), развитие ловушек происходит в отсутствие нематод в условиях относительного недостатка питания или воды. Возможно, в природе эти факторы наряду с морфогенетическими соединениями типа немина регулируют образование ловушек у хищных грибов.
Гибель пойманной нематоды или хотя бы прекращение ее движений часто происходит быстрее, чем мицелий гриба проникнет в ее тело. Предполагают, что хищные грибы образуют токсины, содержащиеся в клейкой жидкости ловушек. Нематотоксины уже обнаружены у многих видов хищных гифомицетов.
Несмотря на большую адаптацию к хищничеству, хищные грибы в течение длительного времени могут развиваться как сапрофиты в почве или на растительных остатках, питаясь различными органическими соединениями и усваивая, как и многие почвенные сапрофиты, минеральные соединения азота. Установлено даже, что лучшее развитие хищных грибов и более активное улавливание ими нематод происходят в присутствии в среде дополнительного энергетического субстрата (сахаров и других соединений). Как и обычные почвенные сапрофиты, эти грибы вполне конкурентоспособны. При внесении в почву они нормально в ней развиваются и сохраняют способность улавливать нематод.
Хищные грибы широко распространены по всей территории земного шара. Они в изобилии встречаются в местах скопления сапрозойных нематод - в почве, на разлагающихся растительных остатках, гниющей древесине, экскрементах животных, на мхах и в водоемах. Их можно обнаружить в ризосфере и на корнях растений. Основное место обитания большинства из них - почва.
Хищные грибы можно рассматривать как экологическую группу почвенных сапрофитов, в процессе эволюции приобретших способность улавливать нематод и питаться дополнительно. Хищничество у грибов появилось, вероятно, в глубокой древности, причем возникло независимо в разных их группах. У гифомицетов этот способ питания должен быть достаточно древнего происхождения, на что указывает их широкое распространение во всех климатических зонах и наличие сложных ловчих приспособлений (Ф. Ф. Сопрунов).
Активное улавливание хищными грибами нематод, способность их существовать в почве, возможность выращивания их в культуре в больших количествах давно привлекали внимание исследователей к этим грибам, как к возможным средствам в биологической борьбе с нематодами. Однако часто в тепличных и мелкоделяночных опытах препараты хищных грибов давали обнадеживающие результаты, но при более широком применении их эффекта не было. Причиной этого была недостаточность знания экологии хищных грибов, что не позволяло предвидеть их поведение в почве и регулировать его в нужном направлении. Для разработки теоретических основ биологического метода борьбы с фитонематодами и другими патогенными нематодами необходимы фундаментальные исследования их экологии, изучение их роли и условий развития в естественных биоценозах.
Хищные грибы представляют большой интерес и как орудия биологической борьбы с нематодами, патогенными для животных. В ряде случаев были получены положительные результаты при использовании препаратов хищных грибов для обеззараживания почвы от личинок анкилостомы, патогенных для человека (Ф. Ф. Сопрунов, Ю. Я. Тендетник), и для борьбы со стронгилятозом лошадей и овец (С. Ф. Шагалин).
|
ПОИСК:
|
При использовании материалов проекта активная ссылка обязательна:
http://gribochek.su/ 'Библиотека о грибах'