Порядок Гифомицеты (Hyphomycetales) (И. И. Сидорова)
Гифомицеты - наиболее обширный и разнообразный в морфологическом и экологическом отношении порядок несовершенных грибов. Он объединяет грибы с одиночными конидиеносцами, а также с конидиеносцами, собран-ными в коремии и спородохии. Деление его на семейства основано на характере аггрегации конидиеносцев, а также на окраске конидиеносцев и конидий. Он подразделяется на 4 семейства: семейство монилиевых (Moniliaceae) с одиночными светлыми конидиеносцами и светлыми конидиями; семейство демациевых (Dematiaceae) с одиночными, но темными конидиеносцами и конидиями; семейство стильбелловых, или коремиалъных (Stilbellaceae), с конидиеносцами, соединенными в коремии; семейство туберкуляриевых (Tuberculariaceae) с конидиеносцами, собранными в спородохии.
Гифомицеты входят в многочисленные экологические группы грибов: почвенные, ксилофильные (обитающие на древесине), паразиты растений, хищные (улавливающие микроскопических животных и питающиеся ими), водные, микофильные (обитающие как паразиты на других грибах), энтомофильные (паразитирующие на насекомых) и др.
Почвенные гифомицеты (М. А. Литвинова)
Важнейшие компоненты почвы - микроскопические растения и животные, обитающие в ней в огромном количестве и принимающие участие в ряде основных превращений веществ, которые совершаются в почве. Можно без преувеличения отметить, что большая часть почвообразовательных процессов, протекающих в природе, в значительной степени обусловлена жизнедеятельностью микроорганизмов из мира растений (бактерий, актиномицетов и микроскопических грибов). В любой почве земного шара обитает масса этих микроорганизмов, в том числе и почвенные микроскопические грибы. В 1 г почвы насчитывают от нескольких тысяч до сотен тысяч зародышей грибов. Однако существующие в настоящее время методы выявления микроорганизмов в почве еще недостаточно совершенны, и поэтому нет точного представления как в отношении абсолютного их числа, так и качественного разнообразия видового состава всех микроскопических грибов, заселяющих различные почвы мира. Многочисленные микологические анализы разных типов почв показали, что сообщества микроскопических грибов в них весьма различны (например, почвы, насыщенные перегноем и более окультуренные, как правило, содержат весьма большое количество грибов, а почвы неудобренные или целинные менее богаты ими). Даже в одном и том же типе почвы появляются различные количества, а также и разные виды микроскопических грибов, обусловленные тем, что пробы почв из разных географических зон отличались различной степенью окультуренности и содержания в них гумуса.
Не все микроскопические грибы, обнаруженные в почве, являются типичными, подлинно почвенными грибами. Типичные почвенные грибы приурочены к постоянному обитанию в почве. В процессе эволюции эти грибы адаптировались к условиям существования в почве за счет использования в ней сравнительно ограниченных источников энергии, в частности углеродных соединений.
В почве также встречаются многочисленные микроскопические фитопатогенные грибы, попавшие в нее вместе с остатками погибших растений.
Микроскопические грибы выполняют в почве весьма разнообразные функции. Гетеротрофное питание грибов обусловливает их непосредственное участие в разложении растительных и животных остатков в почве, т. е. минерализацию разнообразных органических веществ. Грибы разрушают не только простые органические соединения, но и весьма сложные по химическому строению вещества, которые с трудом подвергаются распаду под воздействием других почвенных микроорганизмов (например, бактерий и актиномицетов).
Среди органических веществ, которые составляют основные ингредиенты растительных остатков, попавших в почву и могущих служить источниками энергии для микроскопических грибов, известны крахмал, пектин, клетчатка и лигнин. Почвенные грибы вызывают трансформацию этих сложных органических соединений, в том числе еще ряд других подобных веществ (например, танин, кератин). Расщепление этих продуктов растительного происхождения грибами в почве, безусловно, имеет исключительное значение в общем круговороте веществ в природе. Почвенные грибы также активно участвуют в ферментативном гидролизе белков. У них очень широко распространена протеолитическая способность, обусловливающая интенсивное расщепление белков в почве с выделением аммиака в результате дезаминирования аминокислот. Многие почвенные микроскопические грибы разрушают мочевину благодаря наличию у них фермента уреазы, а также аспарагин под воздействием фермента аспарагиназы. Органические соединения азота для большинства почвенных грибов оказываются прекрасными источниками азота. Наличие в среде аминокислот, пептонов и белков стимулирует развитие многих почвенных микроскопических грибов. Они также хорошо усваивают в почве нитратный и аммиачный азот.
Не менее велика роль микроскопических грибов в разрушении почвенного перегноя. Однако некоторые группы почвенных грибов принимают участие в образовании таких веществ, из которых синтезируется гумус.
Среди почвенных микроскопических грибов имеется большое число и таких, которые способны в процессе жизнедеятельности синтезировать весьма сложные органические соединения - антибиотики, оказывающие губительные воздействия на бактерии и другие микроорганизмы. Антибиотическая активность почвенных микроскопических грибов представляет собой приспособление к условиям среды, обильно заселенной различными микроорганизмами, ведущими между собой и с другими грибами активную борьбу за существование. Например, двенадцать видов пенициллов образуют антибиотик пенициллин. Много продуцентов различных антибиотиков обнаружено среди аспергиллов, триходермы и глиокладиума. Известны грибы, продуцирующие токсигенные вещества и подавляющие рост высших растений.
Касаясь вопроса о роли микроскопических грибов в почвообразовательных процессах, следует заметить, что почвенные грибы, продуцирующие антибиотики, непосредственно принимают участие в синтезе гумуса и гумусоподобных веществ в почве. Химическими исследованиями раскрыта связь между структурой гумуса со строением ряда антибиотических веществ, образованных почвенными микроскопическими грибами. Антибиотические вещества грибного происхождения в большинстве случаев имеют циклическую структуру. В основу химической структуры гумуса входят циклические соединения типа хинонов. В более гумусных почвах преобладают грибы - продуценты антибиотиков с более сложной полициклической структурой. Почвы, в которых обнаружено большое количество микроскопических грибов, продуцирующих антибиотики менее сложной структуры, содержат в гумусе повышенную концентрацию фульвокислот, и, наоборот, в почве, в которой обитает большое число грибов, синтезирующих антибиотики более сложного строения, наблюдают повышенное содержание в гумусе гуминовых кислот. Микроскопические грибы, образуя перегнойные соединения и обволакивая почвенные частицы мицелием, также принимают участие в оструктуривании почвы, что имеет исключительно большое значение в повышении плодородия почвы.
Почвенные микроорганизмы, разрушая и перерабатывая сложные органические вещества г в том числе и перегнойные, переводят их в формы, доступные для питания высших растений. Некоторые продукты обмена веществ грибов также хорошо усваиваются растениями. Особо следует заметить, что в процессе жизнедеятельности почвенные микроскопические грибы синтезируют различные витамины, ферменты и другие физиологически активные вещества, стимулирующие физиологические процессы растений.
Помимо всего перечисленного, процесс разложения отмерших грибов также создает в почве большой запас усвояемой формы пищи для высших растений. Надо иметь в виду, что число поколений микроскопических грибов в почве в зоне корней растений за вегетационный период сменяется от семи до десяти и иногда более раз.
Почвенные грибы и высшие растения находятся в тесной взаимосвязи. Своеобразным и достаточно благоприятным местообитанием для многих почвенных микроскопических грибов является ризосфера, т. е. слой почвы в 2-3 мм, непосредственно примыкающий к корням. Растение пропитывает ризосферный слой почвы своими корневыми выделениями, содержащими различные энергетические вещества, представляющие прекрасный питательный субстрат для развития грибов. Помимо этого, ризосферный слой почвы насыщается корневым опадом, который также хорошо ассимилируется микроскопическими почвенными грибами. Кроме этого, корни растений механическим путем изменяют и разрыхляют структуру почвы, улучшая ее аэрацию. Поэтому в ризосфере обильно развиваются все почвенные микроорганизмы, в том числе и микроскопические грибы.
Количество грибов в ризосфере подчас во много раз превышает число грибов, обитающих в почве вне зоны корней.
В ризосфере растений развиваются сообщества почвенных грибов, которые в той или иной степени по своей структуре видового состава специфичны. Видовой состав сообщества грибов, заселяющих ризосферу, отражает видовой состав микофлоры почвы, так как в ризосфере растений отбирается и формируется группировка микроскопических грибов главным образом из сообщества грибов почвы данной территории. Однако количественные соотношения между отдельными систематическими группами и видами грибов в ризосфере и во флоре грибов, заселяющих почву вне корней, совершенно различны. Например, у большинства травянистых растений в ризосфере бывает усиленное размножение видов родов алътернария (Alternaria) и фузариум (Fusarium), а в почве, расположенной вне корневой зоны растения, их заметно меньше. Исследования ризосферы хлопчатника на юге Украины выявили значительное развитие гриба фузариум оксиспорум (Fusarium oxysporum) и пенициллов (Penicillium) из секции Lanata. Эти грибы вне ризосферной почвы обнаруживают редко. В почвах Каменной степи в ризосфере травянистых растений из целлюлозоразрушающих грибов преимущественно развивается несовершенный гриб фома Маршала (Phoma marchalii), а в почве вне корней его не обнаруживают совсем или встречают редко.
В ризосфере отдельных растений выявляется приуроченность к ним некоторых видов грибов, например у подсолнечника в большом количестве выявлен гриб пеницилл нигриканс (Penicillium nigricans), а в ризосфере озимой пшеницы на этой же почве значительное развитие имел другой гриб - пеницилл коримбиферум (Penicillium corymbiferum). Подобные наблюдения свидетельствуют о специфичности видового состава грибов в ризосфере растений.
Почвенные гифомицеты находятся в сложных взаимоотношениях не только с высшими растениями, но и с почвенными бактериями и акти номицетами. Большая группа почвенных сапрофитных грибов в ризосфере растений способна ограничить развитие различных фитопатогенных микроорганизмов, в том числе паразитных грибов. Эти почвенные грибы используются в качестве биологических мер борьбы с возбудителями болезней сельскохозяйственных растений. Например, почвенный гриб триходерма древесная (Trichoderma lignorum) успешно используется против фитопатогенного гриба вертицилл далие (Verticillium dahliae) - возбудителя увядания (вилта) хлопчатника и других растений.
Многие грибы бывают симбионтами высших растений и образуют с их корнями микоризу.
Несомненно, распространение различных групп почвенных микроскопических грибов в значительной степени обусловлено их физиолого-биохимическими особенностями. Так, мукоровые грибы в качестве азотного питания используют органический, а не минеральный азот. Поэтому эти грибы обильно размножаются на свежих растительных и других остатках, богатых органическим азотом и простыми углеводами. Что касается крахмала и клетчатки, то они как источники углеродного питания очень плохо усваиваются мукоровыми грибами. Сумчатые грибы из рода хетомиум (Chaetomium), наоборот, способны хорошо и энергично усваивать клетчатку. Пенициллы усваивают не только моно- и дисахариды, но и крахмал, а некоторые из них и клетчатку. Аспергиллы, помимо простых источников углерода, ассимилируют и сложные ароматические, в частности непищевые, углеводородные соединения, как, например, некоторые продукты нефти.
На расселение грибов в почве оказывают большое влияние такие факторы, как физические свойства и химический состав почвы, в особенности степень насыщенности почвы органическими веществами, активная кислотность почвы (рН), температура, влажность, обеспеченность кислородом воздуха и, наконец, произрастающие высшие растения в виде целостного фитоценоза, т. е. растительного покрова почвы.
В кислых почвах обычно численность грибов больше, чем в нейтральных, но при этом их видовое разнообразие обычно меньше. По мнению некоторых исследователей, в кислых, а также в щелочных почвах микофлора беднее по видовому составу не из-за угнетающего действия на нее реакции среды, а потому, что в этих условиях многие грибы не выдерживают борьбы за существование с другими микроорганизмами почвы. Часть грибов в этих почвах гибнет под воздействием антагонистов из мира бактерий и актиномицетов. В различных почвенно-климатических зонах при одном и том же значении кислотности (рН) почвы видовой состав грибных сообществ различен. Кроме того, виды грибов, принадлежащие к одному роду, подчас выявляют различное отношение к реакции среды. Например, пеницилл ругулозум (Penicillium rugulosum) и пеницилл лютеум (P. luteum) более часто встречаются в щелочных почвах, а пеницилл фреквентанс (P. frequentans) и пеницилл пулъвиллорум (P. pulvillorum) - в кислых почвах.
Почвенные микроскопические грибы в большинстве являются строгими аэробами, т. е. для своего нормального развития нуждаются в свободном доступе воздуха. По этой причине грибы заселяют главным образом поверхностные слои почвы. В нижерасположенных слоях почвы число грибов обычно невелико. Однако снижение количества грибов в более глубоких слоях почвы обусловлено не только недостаточностью аэрации, но и низким содержанием в них органических веществ. Окультуривание почв приводит к увеличению численности грибов и более глубокому их проникновению в почву. Наибольшее количество почвенных грибов наблюдается в лесной подстилке.
Что касается отношения почвенных грибов к температуре, то большинство из них нормально развивается при 20-25° С, но способны в той или иной степени развиваться при более низкой температуре, примерно до 5° С, и более высокой температуре, доходящей до 30° С. Среди почвенных грибов имеются виды, способные развиваться даже при отрицательной температуре или, наоборот, при весьма повышенной температуре, до 40° С и несколько выше.
Почвенные гифомицеты различают и по отношению их к влажности почвы. Среди них встречаются гигрофилы, развивающиеся при высокой влажности почвы, и есть мезофилы, развивающиеся при средней, т. е. обычной, влажности почвы без капельножидкой влаги, как, например, большинство пенициллов и многие другие. Наконец, известны и ксерофиты, к которым могут быть отнесены многие виды аспергиллов, а также стемфилий (Stemphylium) и другие темноокрашенные гифомицеты.
Проведенные наблюдения в разные годы и в разных районах страны показали, что численный состав грибов в одной и той же почве существенно меняется в течение года и в значительной мере зависит от степени увлажненности и температуры почвы. Например, в дерново-подзолистых почвах Кировской области при теплой и влажной погоде летом наибольшее количество грибов наблюдается в августе; в черноземной почве Каменной степи максимальное количество их наблюдается весной, когда почва хорошо увлажнена и стоит теплая погода.
Повышенное количество микроскопических грибов в почве также обусловлено и другими факторами, например поступлением в почву органического вещества и характером растительного покрова почвы. Наконец, на увеличение численности грибов в почве оказывают влияние различные удобрения и другие сельскохозяйственные мероприятия по окультуриванию почвы.
Богатство почвы гифомицетами во многом зависит от биотических факторов среды. Значительное размножение в почве актиномицетов, бактерий и водорослей подчас подавляет размножение почвенных микроскопических грибов. Однако депрессия у грибов, возникающая под влиянием антагонизма со стороны различных микроорганизмов, заселяющих почву, не у всех видов развивается в одинаковой степени. Некоторые виды грибов успешно противостоят этому губительному воздействию и даже сами способны оказывать отрицательное влияние на развитие бактерий, актиномицетов и других почвенных микроорганизмов. Таким образом, слагающиеся сложные взаимоотношения между почвенными гифомицетами, с одной стороны, и почвенными бактериями, актиномицетами, водорослями и другими микроорганизмами, с другой стороны, предопределяют формирование и развитие в почве на той или иной территории флоры грибов, имеющей определенный и подчас весьма характерный видовой состав.
В северной зоне, в частности в почвах тундры, наиболее крупная группировка микроскопических грибов принадлежит пенициллам. Эти почвы также обильно заселены дрожжевыми и дрожжеподобными организмами. В северных почвах, в том числе почвах тундры, медленно протекают минерализационные процессы, и поэтому почвы насыщаются слаборазложившимися растительными остатками. Именно в таких почвах и развиваются дрожжевые организмы. В почвах тундры почти отсутствуют аспергиллы, редко встречаются грибы родов фузариум (Fusarium) и триходерма (Trichoderma).
Исключительно высокая численность микроскопических грибов обнаружена в почвах тайги. Условия для размножения грибов здесь намного лучше, чем в почвах тундры. Органических веществ, поступающих в почву тайги, значительно больше, чем в почвах тундры; температурные условия также более благоприятны для развития грибов. В этих почвах количество пенициллов в процентном отношении примерно такое же, как и в почвах тундры. Из грибов других родов там много видов триходермы, которые обильно размножаются в почвах с высоким содержанием органических веществ.
Почвы лесолуговой зоны (подзолы и дерново-подзолистые) так же, как и все почвы Севера, содержат большое количество гифомицетов.
В степной зоне и в черноземных целинных почвах число грибов примерно такое же, как и в серых лесных почвах. Количество пенициллов несколько более 50%, но заметно увеличивается число аспергиллов и фузариумов. Дрожжевые организмы встречаются в этих почвах в незначительном количестве. Далее к югу в каштановых почвах число аспергиллов и фузариумов все более и более увеличивается. Что касается общего количества грибов в этих почвах, оно гораздо меньше, чем в почвах Севера.
В зоне пустынных степей в сероземных почвах количество аспергиллов все более возрастает. Эти почвы также обильно заселены фузариумами.
Желтоземы, расположенные в субтропической зоне, особенно богаты грибами, в них так же, как и в других почвах южной зоны, обильно представлены аспергиллы и фузариумы.
В почвах субтропиков обнаружено значительное расселение грибов триходермы. В южной зоне на солонцах, солончаках и такырах растительность скудная, так как условия для жизни растений там исключительно суровые. В этих почвах, как и в других почвах зоны, также обнаружены главным образом аспергиллы. Из пенициллов выявлены виды, принадлежащие главным образом к одной секции моновертициллята (Monoverticillata). В солонцах и такырах до 80-90% видов пенициллов относятся к этой секции. Флора почвенных грибов такыров и такыровидных почв представлена главным образом темнопигментированными гифенными грибами. На такырах, расположенных вблизи Копетдага (Туркмения), из темнопигментированных грибов представители родов стемфилий (Stemphylium) и макроспориум (Macrosporium) составляют 70-75% всех грибов, обнаруженных в этих почвах. В них обильно размножаются пенициллы и слабо развиваются аспергиллы и фузариумы. Однако окультуривание подзолов и дерново-подзолистых почв приводит к некоторому обогащению их грибами рода фузариум. В этих почвах обычно встречаются грибы родов триходерма и мукор.
В лесостепной зоне в серых лесных почвах также преобладают пенициллы, а аспергиллы, как и дрожжи, встречаются редко. Мукоровые грибы и триходермы присутствуют примерно в таких же количествах, что и в почвах лесолуговой зоны. Фузариума обычно мало в серых лесных почвах, но в почве, занятой травянистыми растениями, грибы этого рода встречаются обильно.
Все приведенные примеры достаточно подробно иллюстрируют общие закономерности распределения различных групп почвенных гифомицетов, которые в основном определяются экологическими и географическими факторами окружающей среды и физиологическими особенностями самих грибов. Влияние эколого-географических факторов наглядно выявляется при сравнительном анализе структуры видового состава грибной флоры различных типов почв из разных географических зон. В каждой почвенной климатической зоне существующие сообщества почвенных микромицетов характеризуются довольно специфическим видовым составом и количественными соотношениями между различными систематическими группировками грибов. Влияние растительного покрова также сказывается на флоре почвенных гифомицетов, в особенности на формировании сообществ грибов ризосферы высших растений, но их видовой состав в основном отражает состав гифомицетов, заселяющих почвы данной географической зоны. Наконец, выявилась замена одних видов грибов другими в почвах, находящихся на разных стадиях почвообразовательного процесса, и наличие определенных доминирующих видов грибов в каждом типе почв.
В настоящем обзоре мы не касаемся ряда специализированных группировок гифомицетов, обнаруживаемых в почве, как фитопатогенных паразитных грибов, грибов-микоризообразователей и хищных грибов, приспособившихся к питанию мелкими беспозвоночными животными (амебами, личинками насекомых, почвенными нематодами). Все эти группы грибов должны быть освещены независимо от описания сообщества типичных (истинных) сапрофитных почвенных гифомицетов, имеющих свое место и играющих определенную специфическую роль в почвообразовательных процессах, в общем круговороте веществ в природе и в жизни растений.
Наибольшее количество видов, обитающих в различных почвах, относится к классу несовершенных грибов. Эти грибы наиболее легко выделяются из почвы в культуры, выращиваемые на искусственных питательных средах в лабораторных условиях.
Несовершенные грибы, особенно гифомицеты, наиболее широко распространены в различных почвах мира. Их там обнаруживают не только в виде спор, но и в стадии мицелия. Гифомицеты существуют в почве как за счет использования самих ингредиентов почвы, так и опавших в почву неразложившихся растительных и животных остатков. Многие из этих грибов могут развиваться на живых растениях как патогенные паразиты, вызывая серьезные болезни растений.
Грибы порядка сферопсидных (Sphaeropsidales) представлены в почве единичными видами родов фома (Phoma), кониотирий (Coniothyrium) и хетомелла (Chaetomella). Грибы этих родов на обычные питательные среды из почвы выделяются редко. Грибы рода хетомелла изолируются из почв при посеве почвенных комочков на стерильную фильтровальную бумагу, положенную поверх агаровых пластинок с питательной минерально-азотной средой или на фильтровальную бумагу, увлажненную питательным раствором, состоящим из комплекса минеральных солей, пептона и Сахаров.
Грибы порядка меланкониевых (Melanconiales) в почве представлены одним видом из рода криптомела (Cryptomela) - С. acutispora, обнаруженным в почвах Арабской Республики Египет.
Среди гифомицетов особенно часто в почве встречаются грибы многих родов, в частности ооспора (Oospora), геотрихум (Geotrichum), акремониум (Acremonium), триходерма (Trichoderma), аспергилл (Aspergillus), пеницилл (Penicillium), скопулариопсис (Scopulariopsis), вертицилл (Verticillium), трихотециум (Trichothecium), стемфилий (Stemphylium), фузариум (Fusarium).
Важнейшие почвенные грибы относятся к родам пеницилл, аспергилл и фузариум. Также широко распространены в почвах разных типов следующие гифальные грибы.
У грибов рода триходерма (Trichoderma) мицелий бесцветный или светлый, образующий белые, желтые, чаще зеленые или темно-зеленые колонии. Конидии одноклеточные, шаровидные или эллипсоидные, светлые или бесцветные, часто скученные в небольшие головки.
Эти грибы в большом количестве встречаются в почвах тайги, целинных, лесных и лесолуговой зоны, т. е. в почвах, богатых органическими остатками. Также достаточно обильно они заселяют культурные почвы. В зоне подзолистых почв этих грибов больше, чем в других почвах. Особенно часто их обнаруживают в кислых почвах с низким значением рН (обычно 3,7-5,2).
Триходерма зеленая (Trichoderma viride) и триходерма древесная (Т. lignorum) появляются на чапек-агаре и на сусловом агаре вначале в виде бесцветного мицелия, который быстро разрастается и с возрастом приобретает зеленый цвет. Колонии этих грибов бывают различных оттенков, от лимонно-желто-зеленого цвета до темно-зеленого.
Триходерму зеленую используют для приготовления препаратов: триходермин 1, 2 и 3. Эти препараты изготовляют на основе массового (производственного) размножения этого гриба, с использованием разных растительных отходов и других субстратов (хлебного мякиша, соломенной резки, отходов зерна и хлебных злаков, перегретого торфа). Триходермины применяют для подавления в почве возбудителей болезни льна, корневых гнилей, вертициллезного увядания хлопчатника и других инфекционных болезней сельскохозяйственных растений. Данный гриб также подавляет развитие многоядного паразита - склеротинии Sclerotinia sclerotiorum, вызывающего заболевание склеротиниоз многих растений, в том числе кукурузы и подсолнечника. Гриб образует два известных антибиотика: глиотоксин и виридин, обладающих антибактериальными и антигрибными свойствами (против патогенных бактерий и грибов).
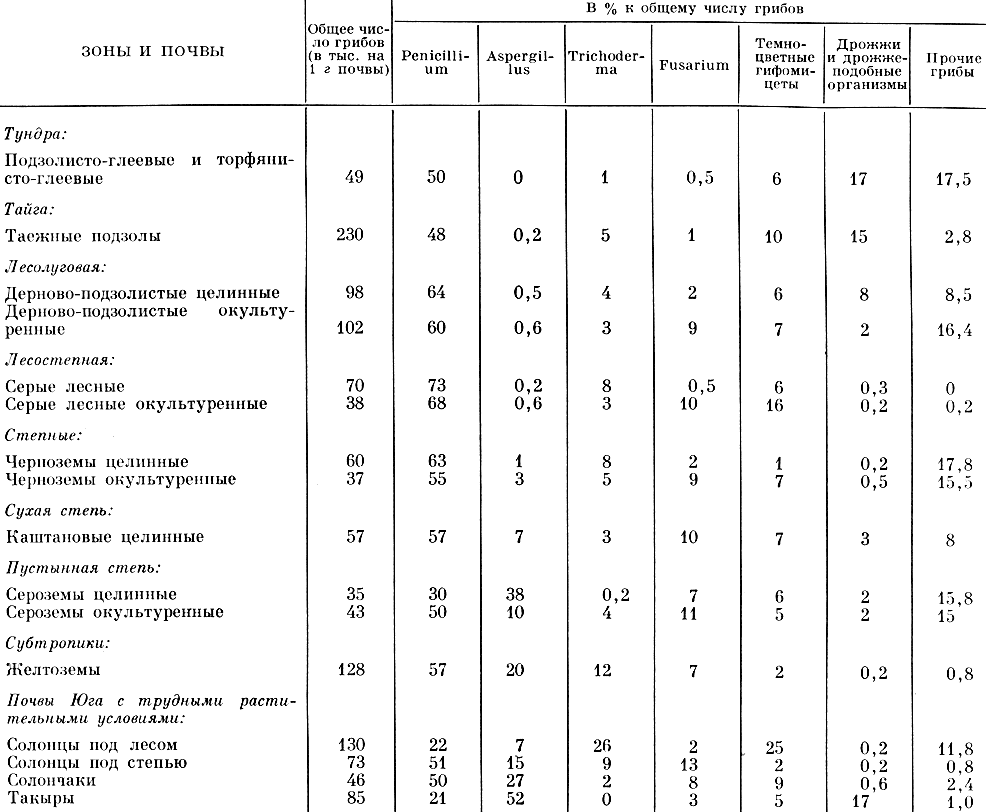
Содержание гифомицетов и дрожжевых организмов в почвах различных типов (по Е. Н. Мишустину, О. Н. Пушкиной и 3. Ф. Тепляковой, 1961)
Триходерма Конинга (Trichoderma koningii) образует колонии на картофельно-сахарозной среде. Вначале они белые, в виде ватообразных клубочков, впоследствии становятся светло-зелеными, но никогда не бывают темно-зелеными. Колонии на чапек-агаре распростертые, пушистые, вначале белые, с возрастом различных оттенков зеленого цвета, но не темно-зеленые.
У грибов рода вертицилл (Verticillium) гифы бесцветные или светлые. Конидиеносцы приподнимающиеся, разветвленные, с ясно выраженной главной осью и отходящими боковыми ветвями; ветви первого порядка большей частью расположены на основной оси мутовчато или супротивно, реже поочередно; ветви второго и последующих порядков - мутовчато; конечные веточки - фиалиды - расположены под острым углом; на фиалиде - по одной конидии. Иногда выделяется слизь, которая склеивает отделяющиеся конидии в ложные головки, быстро распадающиеся. Конидии одноклеточные, шаровидные, обратнояйцевидные, бесцветные или светлые. У некоторых видов образуется темный покоящийся мицелий, микросклероции и хламидоспоры.
Вертициллы - большая группа мутовчатых грибов, которые можно обнаружить при исследовании воздуха, воды и почвы. Наибольшее количество вертициллов встречается в кислых почвах. Среди почвенных вертициллов есть несколько видов, патогенных для растений. Они вызывают увядание растений (вилт). Поражая какое-либо растение, патогенный гриб проникает в сосудистую систему, развивая мицелий в сосудах, проводящих воду, вызывает трахеомикозное заболевание, проявляющееся в увядании растений. В результате этого заболевания растение может потерять всю листву, засыхает и погибает.
Вертицилл кирпично-красный (Verticillium lateritium) имеет красные колонии с белым краем.
Этот вид гриба довольно часто встречается в лесных почвах Западной Сибири. Вертицилл входит в состав комплекса грибов, обусловливающего развитие кагатной гнили свеклы. Гриб поражает корнеплоды во время их хранения в кагатах (в бурах). На зараженных корнях развиваются плесени различного цвета и по мере развития гнили происходит отмирание и разложение ткани корней. Развитию гнили способствуют неправильные условия хранения свеклы.
Грибы рода ауреобазидиум (Aureobasidium) имеют слабо развитый мицелий, вначале бесцветный, позже темный. Гифы состоят из цепочек темных, толстостенных клеток, от которых отпочковываются одноклеточные, овальные или эллипсоидные конидии.
Ауреобазидиум почкующийся (Aureobasidium pullulans). Гифы мицелия состоят из темных с толстой оболочкой клеток.
Грибы рода алътернария (Alternaria), как большинство почвенных темноцветных гифомицетов, наиболее распространены в южных широтах. Особенно часто они встречаются в почве ризосферы многих травянистых растений. Многие из них вызывают заболевания высших растений.
Алътернария тонкая (Alternaria tenuis) имеет колонии дымчато-оливковые или темно-оливковые. Конидиеносцы простые или разветвляющиеся, с перегородками, прямые или слегка коленчато-изогнутые.
Этот вид широко распространен в почвах южных широт. Наибольшее количество гриба обнаружено в черноземной почве, несколько менее - в серой оподзоленной почве и в дерново-подзолистой. Из экстрактов мицелия и культуральной жидкости альтернарии извлекают различные ферменты.
Гриб вызывает заболевание альтернариоз у большого ряда сельскохозяйственных растений (например, ржи, ячменя, кукурузы, риса, гороха, сои).
Ряд грибов рода стахиботрис (Stachybotrys) довольно часто обнаруживается в почвах Севера. Особенно они распространены на природных целлюлозосодержащих субстратах.
Стахиботрис чередующийся (Stachybotrys alternans) имеет мицелий бледно-оливковый, позже оливково-бурый. Развиваясь сапрофитно на мертвых частях растений (стерне, соломе, засохших стеблях различных сорняков), этот почвенный гриб широко распространен в природе. Он принимает участие в разложении растительной клетчатки. В процессе своей жизнедеятельности он образует токсическое вещество, выделяемое им в субстрат. При попадании вместе с соломой в организм лошадей гриб своим токсином вызывает у животных сначала раздражение слизистой оболочки рта и кишечника, а затем, при систематическом потреблении, полное отравление и их гибель. Это заболевание лошадей называют стахиботриотоксикозом.
Группа темноцветных гифомицетов, состоящая главным образом из грибов родов стемфилиум, макроспориум и кладоспориум, широко распространена в почвах южных и умеренных широт по отношению к обильно встречаемым там почвенным грибам из родов пенициллиум и фузариум. Грибы родов стемфилиум, макроспориум, кладоспориум в Туркмении на такырах составляют около 40% всей флоры почвенных грибов. По частоте встречаемости почвенные гифомицеты явно преобладают в южных районах.
Грибы рода кладоспорий (Cladosporium) наиболее распространены в почвах южных широт. Многие виды этого рода - полупаразиты. На многих растениях они образуют темно- или серовато-оливковые бархатистые пятна.
Кладоспорий Траншеля (Cladosporium transchelii) особенно широко распространен в почвах южных районов Украины.
Пенициллы по праву занимают первое место по распространению среди гифомицетов. Естественный резервуар их - почва, причем они, будучи в большинстве видов космополитами, в отличие от аспергиллов, приурочены больше к почвам северных широт.
Как и аспергиллы, они наиболее часто обнаруживаются в виде плесневых налетов, состоящих в основном из конидиеносцев с конидиями, на самых разных субстратах, главным образом растительного происхождения.
Представители этого рода были обнаружены одновременно с аспергиллами благодаря их в общем сходной экологии, широкому распространению и морфологическому сходству.
Мицелий пенициллов в общих чертах не отличается от мицелия аспергиллов. Он бесцветный, многоклетный, ветвящийся. Основное различие между этими двумя близкими родами заключается в строении конидиального аппарата. У пенициллов он более разнообразен и представляет собой в верхней части кисточку различной степени сложности (отсюда его синоним "кистевик"). На основе строения кисточки и некоторых других признаков (морфологических и культуральных) в пределах рода установлены секции, подсекции и серии.
Самые простые конидиеносцы у пенициллов несут на верхнем конце только пучок фиалид, образующих цепочки конидий, развивающихся базипетально, как у аспергиллов. Такие конидиеносцы называют одномутовчатыми или моновертициллятными (секция Monoverticillata, рис. 231). Более сложная кисточка состоит из метул, т. е. более или менее длинных клеток, расположенных на вершине конидиеносца, а на каждой из них находится по пучку, или мутовке, фиалид. При этом метулы могут быть или в виде симметричного пучка (рис. 231), или в небольшом количестве и тогда одна из них как бы продолжает основную ось конидиеносца, а другие располагаются на нем не симметрично (рис. 231). В первом случае они называются сим-метричными (секция Biverticillata-symmetrica), во втором - асимметричными (секция Asymmetrica). Асимметричные конидиеносцы могут иметь еще более сложное строение: метулы тогда отходят от так называемых веточек (рис. 231). И наконец, у немногих видов как веточки, так и метулы могут быть расположены не в один "этаж", а в два, три и больше. Тогда кисточка оказывается как бы многоэтажной, или многомутовчатой (секция Polyverticillata). У некоторых видов конидиеносцы объединяются в пучки - коремии, особенно хорошо развитые в подсекции Asymmetrica-Fasciculata. Когда коремии преобладают в колонии, их можно видеть невооруженным глазом. Иногда они бывают высотой 1 см и больше. Если в колонии коремии слабо выражены, то она имеет мучнистую или зернистую поверхность, чаще всего в краевой зоне.
Детали строения конидиеносцев (гладкие они или шиповатые, бесцветные или окрашенные), размеры их частей могут быть различны в разных сериях и у разных видов, так же как форма, строение оболочки и размеры зрелых конидий (табл. 56).
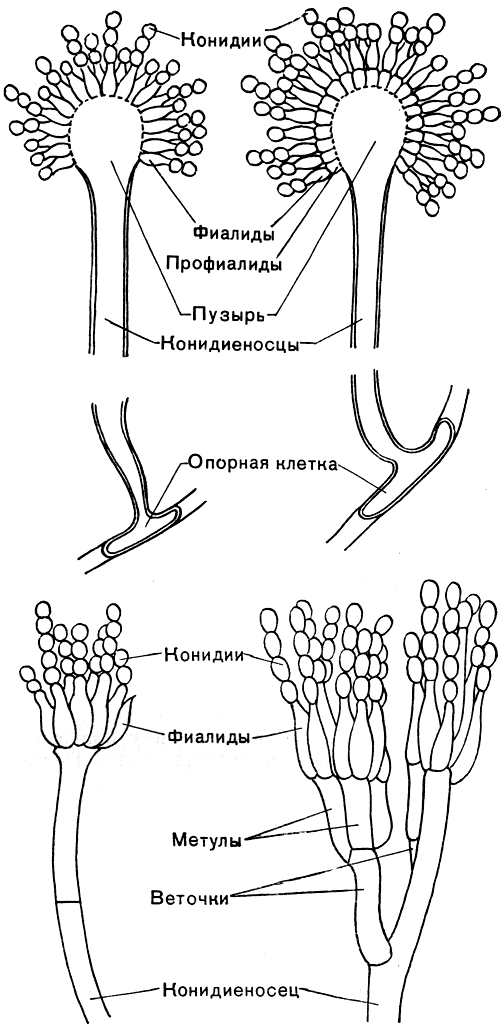
Рис. 231. Строение конидиеносцев: вверху - аепергилл; внизу - пеницилл
Так же как у аспергиллов, у некоторых пенициллов имеется высшее спороношение - сумчатое (половое). Сумки так же развиваются в клейстотециях, похожих на клейстотеции аспергиллов. Эти плодовые тела были впервые изображены в работе О. Брефельдом (1874).
Интересно, что у пенициллов существует та же закономерность, которая отмечена для аспергиллов, а именно: чем проще строение конидиеносного аппарата (кисточки), тем у большего числа видов мы находим клейстотеции. Таким образом, чаще всего они обнаруживаются в секциях Monoverticillata и Biverticillata-Symmetrica. Чем сложнее кисточка, тем меньше в этой группе встречается видов с клейстотециями. Так, в подсекции Asymmetrica-Fasciculata, характеризующейся особенно мощными конидиеносцами, объединенными в коремии, нет ни одного вида с клейтотециями. Из этого можно заключить, что эволюция пенициллов шла в направлении усложнения конидиеносного аппарата, возрастающей продукции конидий и угасания полового размножения. По этому поводу можно высказать некоторые соображения. Так как у пенициллов, как и у аспергиллов, имеется гетерокариозис и парасексуальный цикл, то эти особенности представляют собой ту базу, на основе которой могут возникать новые формы, приспосабливающиеся к разным экологическим условиям и способные завоевать новые жизненные пространства для особей вида и обеспечивать его процветание. В соединении с тем огромным количеством конидий, которые возникают на сложном конидиеносце (оно измеряется десятками тысяч), в то время как в сумках и в клейстотециях в целом количество спор несоизмеримо меньше, общая продукция этих новых форм может быть очень велика. Таким образом, наличие парасексуального цикла и эффективного образования конидий, по существу, обеспечивает грибам ту выгоду, которую другим организмам доставляет половой процесс по сравнению с бесполым или вегетативным размножением.
В колониях многих пенициллов, как у аспергиллов, имеются склероции, служащие, по-видимому, для перенесения неблагоприятных условий.
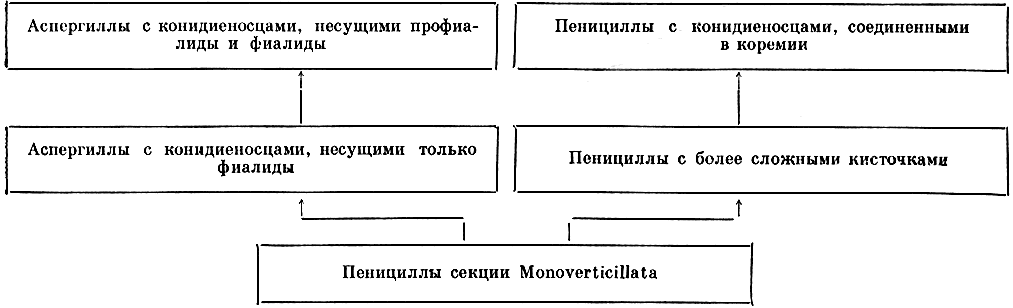
Таким образом, в морфологии, онтогенезе и других особенностях аспергиллов и пенициллов имеется очень много общего, что позволяет предполагать их филогенетическую близость. Некоторые пенициллы из секции Monoverticillata имеют сильно расширенную верхушку конидиеносца, напоминающую вздутие конидиеносца аспергиллов, и, как аспергиллы, встречаются чаще в южных широтах. Поэтому можно представить себе отношения между этими двумя родами и эволюцию в пределах этих родов следующим образом:
Внимание к пенициллам возросло, когда у них впервые была открыта способность образовывать антибиотик пенициллин. Тогда в изучение пенициллов включились ученые самых разнообразных специальностей: бактериологи, фармакологи, медики, химики и т. д. Это вполне понятно, так как открытие пенициллина было одним из выдающихся событий не только в биологии, но и в ряде других областей, особенно в медицине, ветеринарии, фитопатологии, где антибиотики нашли затем самое широкое применение. Именно пенициллин был первым открытым антибиотиком. Широкое признание и применение пенициллина сыграло большую роль в науке, так как ускорило открытие и введение в лечебную практику других антибиотических веществ.
Лечебные свойства плесеней, образуемых колониями пенициллов, были впервые отмечены русскими учеными В. А. Манассеиным и А. Г. Полотебновым еще в 70-х годах прошлого века. Они использовали эти плесени для лечения кожных заболеваний и сифилиса.
В 1928 г. в Англии профессор А. Флеминг обратил внимание на одну из чашек с питательной средой, на которую была посеяна бактерия стафиллококк. Колония бактерии перестала расти под действием попавшей из воздуха и развивавшейся в этой же чашке сине-зеленой плесени. Флеминг выделил гриб в чистую культуру (это оказался Penicillium notatum) и продемонстрировал его способность продуцировать бактериостатическое вещество, которое он назвал пенициллином. Флеминг рекомендовал использовать это вещество и отметил, что его можно применять в медицине. Однако значение пенициллина стало очевидным в полной мере лишь в 1941 г. Флори, Чейн и другие описали методы получения, очистки пенициллина и итоги первых клинических испытаний этого препарата. После этого была намечена программа дальнейших исследований, включавшая поиски более подходящих сред и способов культивирования грибов и получения более продуктивных штаммов. Можно считать, что именно с работ по повышению продуктивности пенициллов началась история научной селекции микроорганизмов.
Еще в 1942-1943 гг. было установлено, что способностью продуцировать большое количество пенициллина обладают также некоторые штаммы другого вида - P. chrysogenum (табл. 57). Активные штаммы были выделены в СССР в 1942 г. профессором З. В. Ермольевой с сотрудниками. Много продуктивных штаммов выделено и за рубежом.
Вначале пенициллин получали, используя штаммы, выделенные из различных природных источников. Это были штаммы P. notaturn и P. chrysogenum. Затем были отобраны изоляты, дававшие более высокий выход пенициллина, сначала в условиях поверхностной, а потом и погруженной культуры в особых чанах - ферментерах. Был получен мутант Q-176, отличающийся еще более высокой продуктивностью, который и использовался для промышленного получения пенициллина. В дальнейшем на основе уже этого штамма были селекционированы еще более активные варианты. Работа по получению активных штаммов ведется непрерывно. Высокопродуктивные штаммы получают преимущественно при помощи сильнодействующих факторов (рентгеновские и ультрафиолетовые лучи, химические мутагены).
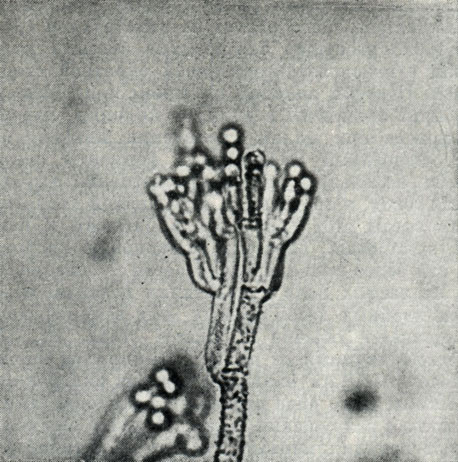
Рис. 232. Конидиеносец пеницилла Penicillium cyclopium
Лечебные свойства пенициллина очень разнообразны. Он действует на гноеродные кокки, гонококки, анаэробные бактерии, вызывающие газовую гангрену, в случаях различных абсцессов, карбункулов, раневых инфекций, остеомиелита, менингита, перитонита, эндокардитов и дает возможность спасти жизнь больных, когда другие лечебные препараты (в частности, сульфамидные) бессильны.
В 1946 г. удалось осуществить синтез пенициллина, который был идентичен природному, полученному биологическим путем. Однако современная пенициллиновая промышленность базируется на биосинтезе, так как он дает возможность массового изготовления дешевого препарата.
Из секции Monoverticillata, представители которой чаще встречаются в более южных районах, наиболее распространен Penicillium frequentans. Он образует на питательной среде широко растущие бархатистые зеленые колонии с красновато-коричневой обратной стороной. Цепочки конидий на одном конидиеносце обычно соединены в длинные колонки, хорошо видимые при малом увеличении микроскопа. P. frequentans продуцирует ферменты пектиназу, используемую для просветления фруктовых соков, и протеиназу. При низкой кислотности среды этот гриб, как и близкий к нему P. spinulosum, образует глюконовую кислоту, а при более высокой кислотности - лимонную.
Из лесных почв и подстилки главным образом хвойных лесов разных мест земного шара выделяется обычно P. thomii (табл. 56, 57), легко отличимый от других пенициллов секции Monoverticillata наличием розовых склероциев. Штаммы этого вида отличаются высокой активностью в разрушении танина, а также они образуют пенициллиновую кислоту - антибиотик, действующий на грамположительные и грамотрицательные бактерии, микобактерии, актиномицеты, на некоторые растения и животных.
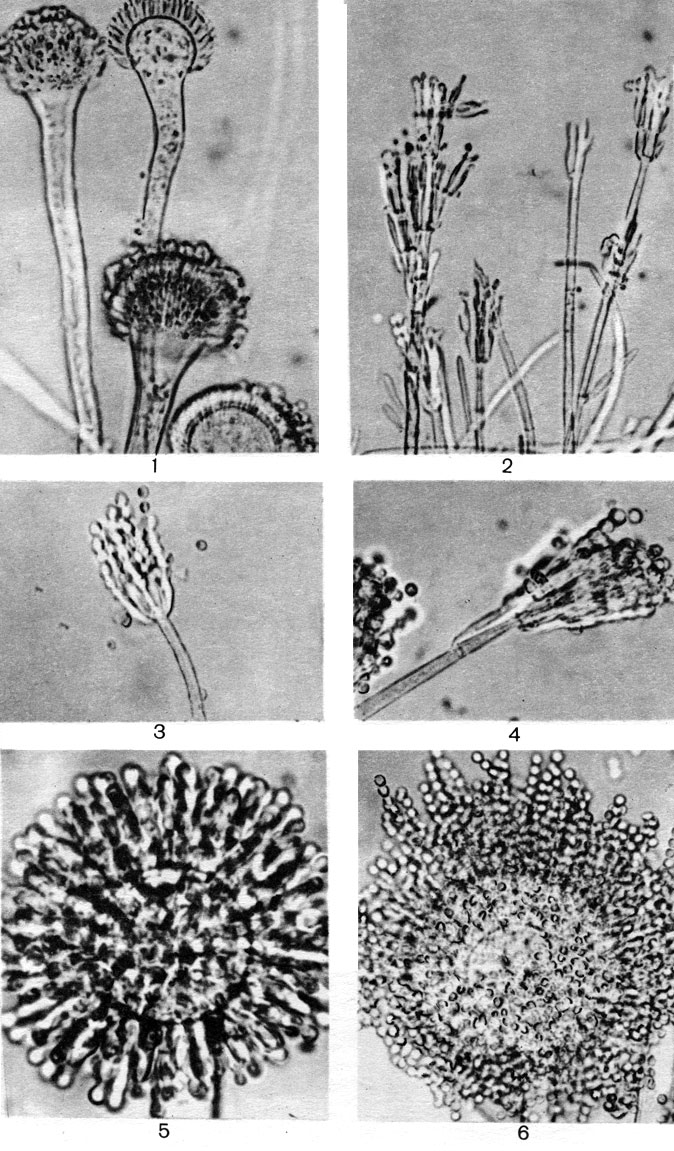
Таблица 56. Конидиеносцы аспергиллов и пенициллов: 1 - аспергилл дымящий (Aspergillus fumigatus); 2 - пеницилл алеющий (Penicillium purpurogenum); 3 - пеницилл Тома (P. thomii); 4 - пеницилл зеленоватый (P. viridicatum); 5, 6 - аспергилл охряный (A. ochraceus)
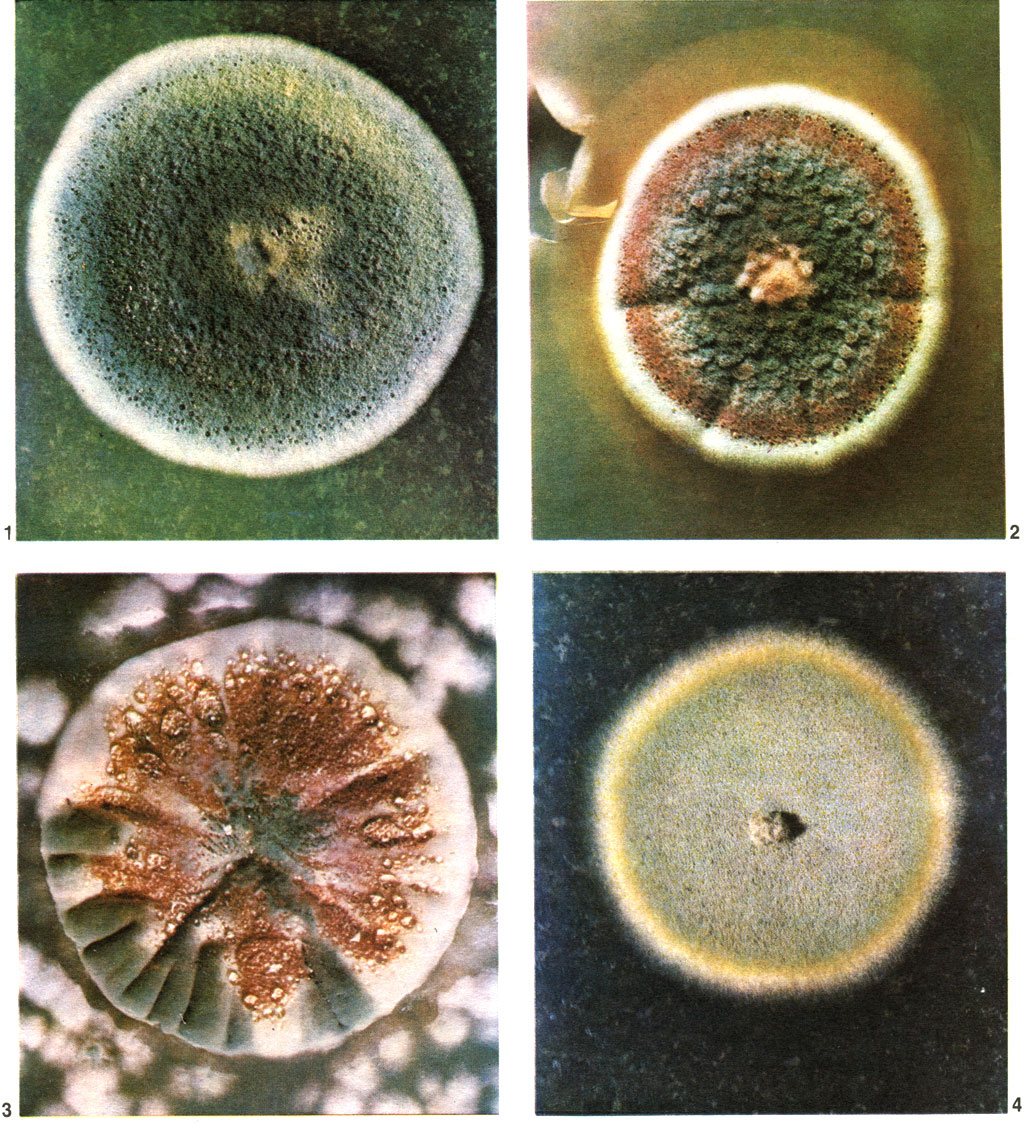
Таблица 57. Колонии пенициллов: 1 - пеницилл отмеченный (Penicillium notatum); 2 - пеницилл с золотистым пигментом (P. chrysogenum); 3 - пеницилл Тома (P. thomii); 4 - пеницилл алеющий (P. purpurogenum)
Многие виды из той же секции Monoverticillata были выделены с предметов военного снаряжения, с оптических инструментов и других материалов в условиях субтропиков и тропиков.
С 1940 г. в странах Азии, особенно в Японии и Китае, известно тяжелое заболевание людей под названием отравления от желтого риса. Оно характеризуется сильным поражением центральной нервной системы, двигательных нервов, расстройством сердечно-сосудистой системы и органов дыхания. Причиной заболевания оказался гриб P. citreo-viride, выделяющий токсин цитреовиридин. В связи с этим было высказано предположение, что при заболевании людей бери-бери наряду с авитаминозом имеет место и острый микотоксикоз.
Не меньшее значение имеют представители секции Biverticillata-symmetrica. Они выделяются из различных почв, из растительных субстратов и промышленных изделий в условиях субтропиков и тропиков.
Многие из грибов этой секции отличаются яркой окраской колоний и выделяют пигменты, диффундирующие в окружающую среду и окрашивающие ее. При развитии этих грибов на бумаге и бумажных изделиях, на книгах, предметах искусства, тентовых покрытиях, обивках автомобилей образуются цветные пятна. Один из основных грибов на бумаге и книгах - P. purpurogenum. Его широко растущие бархатистые желтовато-зеленые колонии обрамлены желтой каймой растущего мицелия, а обратная сторона колонии имеет пурпурно-красную окраску. Красный пигмент выделяется и в окружающую среду.
Иногда P. purpurogenum паразитирует на растениях, в частности на корнях проростков кукурузы, на грибах аспергиллах, поселяется также на личинках некоторых видов комаров. Его споры, находящиеся в воздухе, могут быть причиной аллергических явлений (астмы, сенной лихорадки). Этот гриб вызывает также нередко отомикозы. Он входит в группу почвенных грибов-токсинообразователей и, в частности, угнетает развитие в почве азотфиксирующей бактерии Azotobacter chroococcum.
Особенно большое распространение и значение среди пенициллов имеют представители секции Asymmetrica.
Выше уже говорилось о продуцентах пенициллина - P. chrysogenum и P. notatum. Они встречаются в почве и на различных органических субстратах. Макроскопически их колонии сходны. Они имеют зеленую окраску, и для них, как и для всех видов серии P. chrysogenum, характерно выделение на поверхности колонии эксудата желтого цвета и такого же пигмента в среду (табл. 57).
Можно добавить, что оба эти вида вместе с пенициллином часто образуют эргостерол.
Очень большое значение имеют пенициллы из серии P. roqueforti. Они обитают в почве, но преобладают в группе сыров, характеризующихся "мраморностью". Это сыр "Рокфор", родиной которого является Франция; сыр "Горгонцола" из Северной Италии, сыр "Стилтон" из Англии и др. Всем этим сырам свойственны рыхлая структура, специфический вид (прожилки и пятна голубовато-зеленого цвета) и характерный аромат. Дело в том, что соответствующие культуры грибов используются в определенный момент процесса изготовления сыров. P. roqueforti и родственные виды способны расти в рыхло спрессованном твороге потому, что хорошо переносят пониженное содержание кислорода (в смеси газов, образующихся в пустотах сыра, его содержится меньше 5%). Кроме того, они устойчивы к высокой концентрации соли в кислой среде и образуют при этом липолитические и протеолитические ферменты, воздействующие на жировые и белковые компоненты молока. В настоящее время в процессе изготовления указанных сыров применяют селекционированные штаммы грибов.
Из мягких французских сыров - "Камамбер", "Бри" и др. - выделены P. camamberti и Р. саseicolum. Оба эти вида так давно и настолько адаптировались к своему специфическому субстрату, что из других источников почти не выделяются. В заключительной стадии изготовления сыров "Камамбер" или "Бри" творожную массу помещают для созревания в специальную камеру с температурой 13-14 °С и влажностью 55-60%, воздух которой содержит споры соответствующих грибов. В течение недели вся поверхность сыра покрывается пушистым белым налетом плесени толщиной 1-2 мм. Примерно в течение десяти дней плесневый налет приоб-ретает голубоватый или зеленовато-серый цвет в случае развития P. camamberti или остается белым при преимущественном развитии Р. саseicolum. Масса сыра под воздействием ферментов грибов приобретает сочность, маслянистость, специфические вкус и аромат.
Кроме указанных пенициллов, используемых человеком в столь различных направлениях, среди представителей секции Asymmetriса имеется много вредоносных. Так, большой экономический ущерб причиняют P. digitatum и P. italicum, вызывающие гниение плодов цитрусовых. Часто обе гнили встречаются вместе, но они легко различимы, особенно в начале образования плесневых налетов. P. digitatum - раневой паразит, т. е. в здоровые, неповрежденные плоды его мицелий проникнуть не может. При благоприятных условиях он очень быстро распространяется по поверхности плодов, покрывая их в течение трех-четырех дней зеленовато-оливковым плотным налетом конидий. Пораженные плоды очень быстро сохнут на воздухе, сморщиваются и в заключение покрываются углублениями и мумифицированной коркой грязно-оливково-коричневого цвета ("зеленая гниль" плодов цитрусовых).
P. digitatum выделяет этилен, вызывающий более быстрое созревание здоровых плодов цитрусовых, находящихся поблизости от плодов, пораженных этим грибом.
P. italicum представляет собой сине-зеленую плесень, вызывающую мягкую гниль плодов цитрусовых. Этим грибом чаще поражаются апельсины и грейпфруты, чем лимоны, в то время как P. digitatum развивается с равным успехом на лимонах, апельсинах и грейпфрутах. При интенсивном развитии P. italicum плоды быстро теряют свою форму и покрываются пятнами слизи.
Конидиеносцы P. italicum часто соединяются в коремии, и тогда плесневый налет приобретает зернистость. Оба гриба имеют приятный ароматический запах.
В почве и на различных субстратах (зерне, хлебе, промышленных товарах и т. п.) часто встречается P. expansum (табл. 58).Но особенно известен он как причина быстро развивающейся мягкой коричневой гнили яблок. Потери яблок от этого гриба при хранении составляют иногда 85-90%. Конидиеносцы этого вида также образуют коремии. Массы спор его, присутствующие в воздухе, могут вызывать аллергические заболевания.
Некоторые виды коремиальных пенициллов приносят большой вред цветоводству. P. corymbiferum выделяется с луковиц тюльпанов в Голландии, гиацинтов и нарциссов в Дании. Установлена также патогенность P. gladioli для луковиц гладиолусов и, по-видимому, для других растений, имеющих луковицы или мясистые корни.
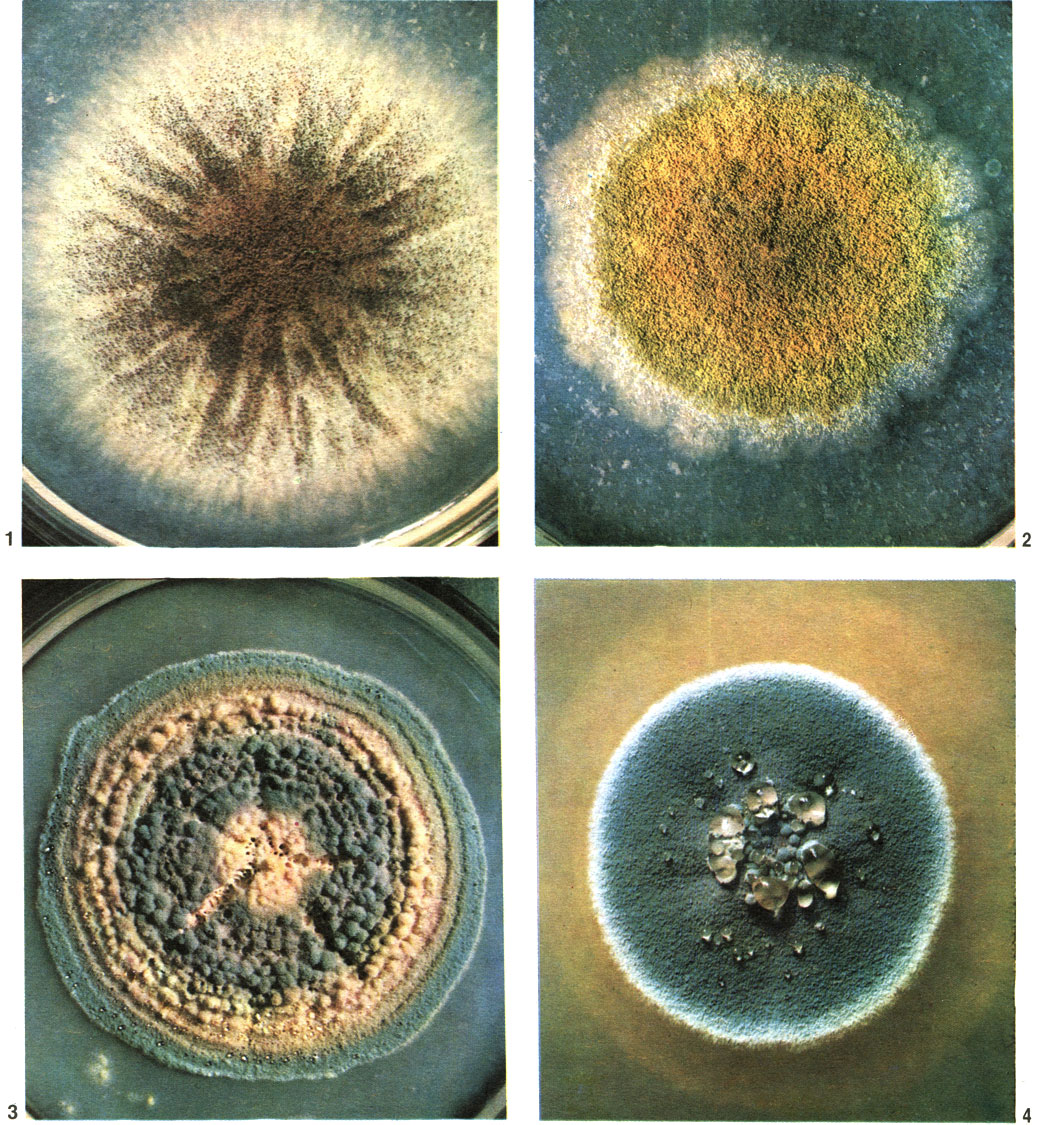
Таблица 58. Колонии пенициллов и аспергиллов: 1 - аспергилл черный (Aspergillus niger); 2 - аспергилл желтый (A. flavus); 3 - пеницилл распростертый (Penicillium expansum); 4 - пеницилл циклопический (P. cyclopium)
Большое значение из коремиальных грибов имеют пенициллы из серии P. cyclopium. Они широко распространены в почве и на органических субстратах, часто выделяются с зерна и зерновых продуктов, с промышленных товаров в разных зонах земного шара и отличаются высокой и разнообразной активностью.
P. cyclopium (рис. 232) принадлежит к одним из самых сильных токсинообразователей в почве.
Некоторые пенициллы секции Asymmetrica (P. nigricans) образуют антигрибной антибиотик гризеофульвин, который показал хорошие результаты в борьбе с некоторыми болезнями растений. Его можно использовать для борьбы с грибами, вызывающими заболевания кожи и волосяных луковиц у людей и животных.
По-видимому, наиболее процветающими в природных условиях оказываются представители секции Asymmetrica. Они имеют более широкую экологическую амплитуду, чем другие пенициллы, лучше других переносят пониженную температуру (P. puberulum, например, может образовывать плесневые налеты на мясе в холодильниках) и относительно меньшее содержание кислорода. Многие из них встречаются в почве не только в поверхностных слоях, но и на значительной глубине, особенно коремиальные формы. Для некоторых видов, как, например, для P. chrysogenum, установлены очень широкие температурные границы (от -4 до +33 СС).
Имея широкий набор ферментов, пенициллы заселяют различные субстраты и принимают самое активное участие в аэробном разрушении растительных остатков.
Использование продуктов обмена веществ пенициллов далеко не исчерпано, и дальнейшее изучение, без сомнения, откроет новые возможности их применения в различных отраслях народного хозяйства.
Род Аспергилл (Aspergillus) (Т. П. Сизова)
Грибы из рода аспергилл, описанного впервые в 1729 г. итальянским микологом П. Микели,- одни из наиболее распространенных гифомицетов. Естественное местообитание их - верхние горизонты почвы, особенно в южных широтах. Но чаще всего их обнаруживают на различных продуктах, главным образом растительного происхождения, где колонии их образуют плесневые налеты разного цвета, особенно часты голубовато-зеленые, реже других цветов. Колонии аспергиллов появляются на хлебе, хранящемся при повышенной влажности, на варенье, влажных обоях, изделиях из кожи и т. п. Следовательно, большинство видов аспергиллов - сапрофиты. Но в этом роде имеются и паразиты животных и человека.
Вегетативное тело аспергиллов - многоклетный, очень ветвистый мицелий, пронизывающий субстрат. Клетки мицелия многоядерны. Иногда развивается и обильный воздушный мицелий. У большинства аспергиллов плесневый налет состоит из конидиеносцев с конидиями. Конидиеносцы отходят вверх от особых клеток мицелия - опорных клеток (рис. 231). У разных видов конидиеносцы имеют различные размеры, могут представлять собой одну клетку или, реже, иметь перегородки, у немногих ветвятся. У большинства аспергиллов конидиеносцы бесцветны, как и гифы мицелия, а у некоторых (например, у представителей групп A. nidulans, A. ochraceus) они коричневатые или желтоватые. Оболочки их у большинства гладкие, у немногих (группа A. ochraceus, A. effusus из группы A. nidulans) шиповатые. Верхняя часть конидиеносца вздувается, образуя пузырь, у большинства округлый, у отдельных видов в различной степени вытянутый. На пузыре или радиально, или только в верхней его части размещаются флажковидные клетки - фиалиды, из узкого горлышка которых выходят одна за другой, располагаясь в цепочку, одноклетные конидии. Непосредственно у отверстия наверху фиалиды конидии имеют тонкую оболочку, часто бесцветны или бледно окрашены, и окончательное созревание и оформление их происходит постепенно. Чем выше по цепочке, тем конидии крупнее, интенсивнее окрашены и более зрелы (табл. 56). Зрелые конидии имеют определенную форму и окраску, у многих аспергиллов становятся при созревании шиповатыми или бугорчатыми. Окраска массы конидий как раз и придает тот или иной цвет плесневому налету, т. е. колонии.
У некоторых аспергиллов, находящихся, по-видимому, на более высокой ступени эволюционного развития, увеличение количества образующихся конидий происходит в результате того, что на пузыре конидиеносца возникает серия клеток (профиалид), на которых сидят пучками, или мутовками, фиалиды (рис. 231). При малом увеличении микроскопа верхушка конидиеносца аспергилла, несущая цепочки конидий, внешне очень похожа на наконечник лейки, из отверстий которого льются струйки воды. Поэтому русское название аспергилла - леечный гриб. Однако точный перевод аспергилла будет "косматая голова", что также очень хорошо согласуется с впечатлением, производимым при рассматривании конидиеносцев с цепочками конидий.
По мере созревания конидии отваливаются, переносятся на новые места и прорастают при благоприятных условиях, образуя мицелий.
При помощи конидий, т. е. бесполым путем, размножается большинство аспергиллов. Однако некоторым из них свойственно и высшее спороношение - сумчатое, т. е. половое. В колониях таких видов бывают заметны невооруженным глазом маленькие шарики, у большинства желтого цвета. Это плодовые тела - клейстотеции. Они были впервые обнаружены Г. Линком (1809) в гербарном материале.
Клейстотеции сохранились главным образом у тех аспергиллов, у которых конидиеносцы имеют более простое строение: не несут на своем пузыре тех дополнительных элементов (профиалид), которые характерны для видов, утративших сумчатое спороношение.
В настоящее время сумчатая стадия известна почти у всех видов группы A. glaucus, у всех видов серии A. fischeri из группы A. fumigatus, у многих видов группы A. nidulans и у некоторых видов групп A. ornatus, A cremeus и A. ochraceus. Эти грибы имеют фиалиды, образующиеся непосредственно на пузыре конидиеносца (без профиалид).
На клейстотеции похожи склероции, образующиеся у многих видов аспергиллов из групп A. candidus, A. niger, A. flavus и A. ochraceus. Но, в отличие от клейстотециев, они представляют собой простое сплетение гиф мицелия и внутри нет сумок. Можно предположить, что склероции - это клейстотеции, утратившие способность формировать аскоспоры. Подтверждением такого предположения служит образование внутри склероциев сумчатого спороношения, например у A. alliaceus.
Склероции особенно обильны при высоком содержании в среде нитратов и сахарозы и при температуре 20-25°С.
Иногда за клейстотеции или склероции можно принять сходные по внешнему облику образования, наблюдающиеся в колониях некоторых групп (A. nidulans, A. versicolor, A. ustus). Это скопления шаровидных или извитых клеток с такими толстыми оболочками, что полость этих клеток почти полностью облитерируется. Иногда их принимают за хламидоспоры. Действительно, у A. nidulans прослежено прорастание этих клеток в мицелий. Однако истинная природа толстостенных клеток не установлена.
У аспергиллов, так же как у пенициллов и многих других грибов, имеются дополнительные механизмы образования новых форм, кроме полового процесса. Первое сообщение о гетерокариозе у A. niger (табл. 58) и A. nidulans, было опубликовано в 1940 г. Г. X. Госсопом с сотрудниками. Парасексуальный процесс, существенный момент в котором представляет гетерокариоз, был также впервые продемонстрирован на A. nidulans в 1952 г. Ж. Е. Ропером, а затем на A. niger Л. Понтекорво и др. С тех пор он был всесторонне изучен у многих других видов и родов грибов.
То, что именно аспергиллы были подвергнуты такому усиленному изучению в указанном плане, отнюдь не было случайным. Примерно с середины XIX в. аспергиллы уже привлекали к себе внимание как активные агенты процессов разрушения самых разнообразных материалов, как возбудители заболеваний и причина токсикозов животных и человека, как продуценты различных ферментов и других ценных продуктов обмена веществ, с успехом используемых в ряде отраслей промышленности. Кроме того, они хорошо растут в лабораторных условиях, почему и стали одним из излюбленных объектов исследований. Достаточно сказать, что между 1891 и 1928 гг. было опубликовано более 2000 работ по аспергиллам, посвященных главным образом физиологии, биохимии и генетике видов этих грибов. Изучение их продолжается и в настоящее время. Особенно активно работают с аспергиллами в лабораториях с 1940 г., когда их стали использовать как вообще весьма удобные модели в исследованиях генетических зако-номерностей, различных физиологических процессов, путей обмена веществ и т. п.
Некоторые группы и виды аспергиллов широко распространены в природе и имеют, как уже было сказано, весьма существенное практическое значение. Это в первую очередь представители группы A. niger, наиболее активно используемые в промышленности и в лабораторных исследованиях. Только видам этой группы начиная с 1945 г. было посвящено свыше 1000 научных статей! Они выделяются из почвы в разных странах и на разных континентах, т. е. являются космополитами. Они развиваются на зерне во время его хранения, на плодах, овощах, хлопчатобумажных изделиях, коже и материалах, богатых белками.
Колонии видов группы A. niger коричневые, шоколадные или черные (табл. 58). Конидиеносцы несут профиалиды с мутовками фиалид.
Виды группы A. niger отличаются многообразной биохимической активностью. Они вырабатывают разнообразные ферменты - амилолитические, протеиназы, пектиназу, липазу, глюкозооксидазу, ферменты, разрушающие роговое вещество, хитин и др. Уже в 1787 г. было описано "сбраживание" галловых орешков при помощи A. niger, вырабатывающего фермент таназу, с образованием галловой кислоты, кото-рая используется для производства ряда красок, чернил, некоторых фармацевтических препаратов. В настоящее время при помощи пектолитических ферментов A. niger и A. foetidus в ряде стран производят осветление фруктовых соков и вин. В Японии пектолитические ферменты A. niger используют для расщепления на волокна стеблей растений (рами). Использование ферментов A. awamori позволяет получать кристаллическую глюкозу из крахмала. Из инулина под действием фермента A. niger можно получать фруктозу, и т. д.
Широкое применение получила способность штаммов A. niger и других видов этой группы к образованию лимонной, щавелевой, глюконовой, фумаровой кислот. В СССР и в других странах для производства лимонной кислоты используются штаммы этого вида. Штаммы группы A. niger способны синтезировать витамины биотин, тиамин, рибофлавин и др.
Что касается антибиотических веществ, то их продуцируется ограниченное количество и практическое использование их невелико.
Применение штаммов A. niger в различных лабораторных исследованиях многообразно. В свое время этот вид образно назвали "биохимической лягушкой" (Л. И. Курсанов). Так как они очень чувствительны к минеральным источникам питания, возможно использовать специально отобранные штаммы для определения дефицита некоторых элементов в почве (фосфора, калия, меди и др.) и витаминов, что оказалось значительно проще, точнее и быстрее, чем химические анализы.
В ряде тропических стран отмечены заболевания растений, вызываемые штаммами A. niger, например проростков арахиса, хлопчатника, сорго в Восточной Африке (Судан), Индии. Гриб A. carbonarius описывают как причину гнили винограда в Индии.
Штаммы A. niger, выделенные из заплесневелых кормов, оказались токсичными для животных. Известны случаи отомикозов, легочных аспергиллезов, бронхопневмонии, мицетом конечностей, причиной которых был A. niger.
Не меньшее значение имеют грибы группы A. flavus-oryzae. Для них характерна желтовато-зеленая окраска колоний. Конидиеносцы у некоторых видов несут на вздутии только фиалиды, а у других и профиалиды. Эти грибы встречаются в почве и на самых разнообразных субстратах: растительных остатках, фураже, пищевых продуктах, растительных маслах, пластических массах и других материалах. A. flavus может расти даже на таких, казалось бы, мало подходящих субстратах, как воск, парафин. Грибы этой группы - главные компоненты сообщества плесневых грибов, развивающихся на зерне и семенах, главным образом на рисе, горохе, соевых бобах, арахисе, в плохо проветриваемых хранилищах, даже при 18% влажности. Разнообразие заселяемых субстратов объясняется тем, что у видов этой группы имеется особенно богатый набор ферментов. Они продуцируют амилазу, протеиназы, липазы, пектиназы (пектазу и протопектиназу), целлюлазу и др. Именно поэтому A. oryzae и родственные виды используются на Востоке для пищевых и других целей в течение не одного столетия. Спиртовая промышленность Японии и других стран Востока целиком основана на ферментативных свойствах грибов этой группы. Ферменты A. oryzae ("така-койи") гидролизируют крахмал рисовых зерен при изготовлении рисовой водки - сакэ. Для получения промышленного спирта в Японии в настоящее время с успехом используются и другие источники крахмала, например батат.
При приготовлении продуктов питания из сои и других субстратов, богатых белками, на Востоке (главным образом в Японии) используют уже в течение столетия протеолитические ферменты A. flavus, A. oryzae, A. parasiticus. В производстве китайского и японского соевого соуса ферментативным способом применяют особую закваску, представляющую собой культуру A. oryzae на вареной сое или перловой крупе. Во Вьетнаме ферментативный соево-рисовый соус "тыонг" служит обязательным повседневным пищевым продуктом населения. При приготовлении его протеолитические и амилолитические ферменты плесневых грибов, и лучше других A. oryzae, гидролизируют крахмал и белки сои и риса. В современных условиях ферментные препараты A. flavus, A. oryzae и A. parasiticus с успехом применяют для очистки кож от волос и размягчения кож, для удаления серебра из старых пленок и пластинок, в текстильной промышленности. В СССР также используют ферменты грибов этой группы, например, для изготовления ферментных препаратов, в производстве спиртов и для других целей.
В настоящее время много внимания уделяют селекции штаммов с высокой α-амилазной, мальтазной и протеазной активностью.
Этим не исчерпывается перечень полезных метаболитов. Способность грибов этой группы образовывать коевую кислоту была установлена в начале нашего столетия. Коевую кислоту вырабатывают A. flavus, A. tamarii, A. parasiticus, A. effusus. Они образуют и другие кислоты (β-нитропропионовую, α-кетоглутаровую, лимонную, яблочную, молочную, фумаровую и др.), а также витамины (рибофлавин, тиамин, пантотеновую кислоту, инозитол, биотин, пиридоксин, В12, С, К3 и др.) и стероиды (тестостерон, тестололактон).
Многообразны проявления паразитных свойств A. flavus и других видов группы по отношению к растениям, насекомым, позвоночным животным и человеку. В частности, от A. Flavus нередко страдают проростки хлопчатника. Он вызывает паралич пчел, заболевания шелковичных червей, термитов и других животных, отомикозы у человека. A. parasiticus вызывает легочные заболевания у голубей, цыплят, индюшек, диких птиц, содержащихся в неволе, у лошадей. У людей констатированы случаи инфекции легких и, кроме того, различные внутриполостные поражения, эндокардиты, грануломатоз головного мозга, причиной которых бывает A. parasiticus.
В последние годы A. flavus получил печальную известность как продуцент одного чрезвычайно вредоносного токсического вещества. Драматическая история открытия этого токсина начинается с 1960 г., когда в Англии появилось загадочное заболевание индюшек, при котором они погибали в большом количестве без каких-либо видимых признаков уже известных заболеваний. На одной из ферм в течение двух недель из 1000 молодых индюшек и индюшат погибло 800. А немногим более чем за три месяца на фермах, специализировавшихся на разведении индюшек, их количество уменьшилось на сто с лишним тысяч.
В расследовании причин заболевания приняли участие лаборатория судебной экспертизы Скотленд-Ярда, Общество охраны животных и многие другие организации. Сразу же возникло подозрение на отравление птичьего корма, который и стали проверять в первую очередь на содержание опасных химических веществ, ядов и различных болезнетворных организмов. Все эти испытания дали отрицательные результаты. Тогда обратили внимание на то, что все вспышки болезни "икс", как ее назвали, происходили на фермах, которым поставляли корм две фабрики, добавлявшие, как выяснилось, в корм муку из земляных орехов (арахиса), вывезенных из Бразилии.
Стало известно, что в Кении погибали подобным же образом утята, которым давали корм с добавкой земляных орехов, привезенных из Уганды. И тогда научные сотрудники Лондонского института тропических культур установили, что токсин в больших количествах вырабатывается грибом A. flavus, быстро развивающимся на земляных орехах в условиях высокой влажности и умеренно высокой температуры тропиков и субтропиков. Токсическое вещество назвали афлатоксин (от начальных букв названия гриба). Позже выяснилось, что это не одно вещество, а целый комплекс (B1, В2, G1, G2). Впоследствии был установлен и канцерогенный характер токсина.
Это открытие имеет, помимо медицинского, большое экономическое значение, поскольку во многих районах мира возделывание земляного ореха - одна из основных отраслей хозяйства. Некоторые из стран Африки почти целиком зависят от производства и экспорта земляного ореха и получаемого из него масла. В связи с этим обеспечение надлежащего хранения этого продукта приобретает первостепенное значение, тем более, что последующие опыты показали, во-первых, что афлатоксин оказывает вредное действие (токсическое и канцерогенное) на широкий круг животных, не исключая, по-видимому, и человека, а во-вторых, что и многие другие грибы, развивающиеся на хранящихся продуктах, особенно на земляных орехах, льняном и хлопковом семенах, ливере, рыбе и т. п., тоже продуцируют токсические вещества и могут стать причиной тяжелых заболеваний.
Нельзя не остановиться подробнее на представителе другой группы аспергиллов (группы A. fumigatus) - A. fumigatus, который часто может быть причиной тяжелых заболеваний животных и человека. У этого гриба известно два типа колоний: пушистые, в которых хорошо развит воздушный белый мицелий и слабо представлено конидиальное спороношение, придаю-щее колонии нежно-голубоватый оттенок, и бархатистые - с мицелием в субстрате и обильным конидиальным спороношением, имеющим густую голубовато-зеленую окраску. При рассматривании колонии под малым увеличением микроскопа видно, что цепочки конидий на каждом конидиеносце образуют все вместе плотную колонку. На поверхности вздутия конидиеносца имеются только фиалиды, покрывающие главным образом его верхнюю часть.
A. fumigatus встречается в почве, развивается на фураже, на различных семенах и зерновых продуктах при хранении, на шерсти, хлопке и т. п. Разрушение этих материалов обычно сопровождается значительным повышением температуры. В компостах, например, при высокой температуре (до +50°С) A. fumigatus составляет примерно 70% от общего количества обнаруживаемых там грибов. Можно сказать, что это самые термофильные грибы среди аспергиллов. Вероятно, как раз этим свойством объясняется то, что A. fumigatus чаще других аспергиллов встречается как паразит животных и человека. Именно он основной паразит домашних и диких птиц, у которых поражает дыхательные пути. У людей этот гриб вызывает легочный аспергиллез, хроническую эмфизему легких и аллергии с симптомами ангины, но чаще бывает причиной тяжелых отомикозов.
A. fumigatus образует токсин, оказывающий гемолитическое и антигенное действие. Экстракт из мицелия этого вида раздражающе действует на кожу и почки.
Аспергилл дымящий (A. fumigatus) продуцирует антибиотик фумагиллин, активный против стафилококкового бактериофага и, главное, обладающий лечебными свойствами против амебной дизентерии, нозематоза пчел, активно тормозящий развитие ряда перевивных опухолей. В лаборатории антибиотиков МГУ под руководством З. Э. Беккер и А. Б. Силаева был получен отечественный препарат кристаллического фумагиллина.
Из мицелия A. fumigatus выделен алкалоид фестуклавин, используемый в медицине и ранее получаемый только из спорыньи (Claviceps purpurea), и два новых алкалоида - фумигаклавины А и В.
В последнее время грибу A. fumigatus приписывают активную роль в разрушении хитиновых веществ в почве. Широкое распространение штаммов этого гриба в различных условиях, многообразная биохимическая деятельность, свойственная им, особенно в условиях повышенных температур, большая стойкость (их мицелий и споры могут сохранять жизнеспособность после пребывания в течение 6-8 мин, в 60%-ном спирте) обусловливают возрастающий интерес к изучению этой группы.
Широко растущие темно-зеленые бархатистые колонии A. nidulans (табл. 58) из группы того же названия также довольно часто обнаруживаются на различных материалах растительного происхождения. Иногда он выделяется из почвы и из тканей животных и человека, развиваясь главным образом в легких и воздушных мешках цыплят, легких лошадей, в ушных проходах животных и человека.
Многие виды группы A. nidulans образуют ярко-желтые клейстотеции, резко выделяющиеся на темном фоне колонии. В зависимости от соотношения количества клейстотециев и конидиального спороношения окраска колоний меняется от темно-зеленой, если преобладает конидиальное спороношение, до ярко-желтой в случае массового развития клейстотециев. Аскоспоры видов этой группы пурпурно-красные. В колониях обычно присутствуют и толстостенные клетки в виде крупных желтых скоплений.
Особо следует указать, что группа A. nidulans занимает второе место после видов рода нейроспора (Neurospora) в генетике грибов. Получено большое количество мутантов, изучают механизм образования митотических рекомбинаций и факторов, определяющих эти процессы, образование гетерокарионов, цитоплазматическую наследственность и т. д.
A. nidulans и полученные из диких штаммов мутанты используют для изучения углеводного, азотного и других типов обмена.
Очень интересна и своеобразна группа A. versicolor. Для ее представителей характерно образование узкорастущих, выпуклых, плотных колоний, различных желтовато-зеленых или синеватых оттенков в конидиальной зоне, часто с розовыми тонами в зоне мицелиального роста. С обратной стороны колонии бывают ярко-красные или вишнево-красные. Такого же цвета пигмент выделяется в среду вокруг колоний.
Наиболее распространены виды A. versicolor и A. sydowii. Они выделяются из почв, особенно южного происхождения, развиваются на зерне, сыре и других продуктах, на различных материалах разного оборудования, автопокрытиях и обивке машин, особенно в условиях тропиков и субтропиков. Выделяя пигмент, они портят некоторые изделия, образуя на них ярко окрашенные пятна в тех участках, где развивались колонии.
Гриб A. versicolor преобладал на пшенице, импортируемой в Италию из Аргентины, США, Сирии и других стран. В музеях многих городов этот гриб был обнаружен на станковой и монументальной живописи. В библиотеках, где 80% повреждений переплетов из дерматина, коленкора или кожи вызываются аспергиллами, он встречается чаще других грибов. Борьба с такого рода повреждениями затрудняется тем, что гриб проявляет устойчивость к большинству антисептиков. Благодаря этому свойству его широко используют при изучении средств, употребляемых для защиты от грибов различных промышленных изделий (например, оптических инструментов, пластических масс и пр.), для оценки устойчивости бумаги к плесневению, для определения эффективности фумигации и других видов обработки почв вегетационных домиков и т. д.
Мы кратко осветили значение только некоторых видов аспергиллов. Однако сказанного достаточно, чтобы представить себе ту огромную роль в природе и хозяйственной деятельности человека, которую они выполняют. Свойственная грибам этого рода широкая экологическая амплитуда дает возможность для развития тех или иных видов при различных условиях окружающей среды. Например, известно, что большинство грибов, в том числе и аспергиллы, активно растут на органических материалах при низких значениях рН. Оказывается, некоторые из них (A. clavatus) не только способны переносить сильное подщелачивание среды, но и сами вызывают это подщелачивание, при котором большинство грибов развиваться не могут. Благодаря этой особенности виды группы A. clavatus часто образуют плесневые налеты на животных тканях.
Многим аспергиллам свойствен ксерофитизм. Так, некоторые представители самой большой по количеству видов группы A. glaucus (A. repens, A, ruber, A. amstelodami) и группы A. ustus хорошо развиваются на зерне и других продуктах, на текстильных изделиях, изоляционных материалах и иных субстратах при низком уровне влажности.
Широкий набор ферментов позволяет аспергиллам осваивать самые разнообразные субстраты. Антибиотические вещества, продуцируемые ими, обеспечивают успешную борьбу с возможными конкурентами. Это создает предпосылки для использования некоторых видов при разработке мер биологической борьбы с фитопатогенными организмами.
Несовершенные грибы-сапрофиты на древесине и растительных остатках (Т. П. Сизова)
Жизнь каждого растения, как и животного, имеет начало и конец. В любом лесу наряду с живыми деревьями, кустарниками, травами есть бурелом, валежник, отломившиеся и упавшие на землю сучья и ветви. Везде бывает более или менее мощный слой подстилки, состоящей главным образом из опавших листьев, хвои ит. п. В полях, на лугах, в садах после периода вегетации растений также накапливается много растительных остатков. Вся эта масса органического вещества разлагается в основном под воздействием грибов, превращаясь в более простые соединения, а затем в почву. Таким образом грибы выполняют важную функцию в общем круговороте веществ в природе. Основная роль при этом принадлежит высшим базидиальным и сумчатым грибам. Но некоторые несовершенные грибы также принимают в этом участие.
Биологическое разложение древесины или других растительных остатков представляет собой очень сложный процесс, осуществляемый на разных этапах разными организмами и зависящий от особенностей разлагающегося субстрата и от тех условий, в которых он протекает. На основании оригинальных исследований В. Я. Частухина и последующих работ отечественных и зарубежных ученых можно представить себе, что разложение древесных остатков в лесу происходит по следующей схеме.
На свежеупавшем дереве прежде всего поселяются типичные деревоокрашивающие грибы, использующие главным образом содержимое клеток. Таковы грибы из родов кладоспорий, альтернария и некоторые другие, вызывающие появление на древесине серовато-синих, коричневых и черных окрасок; фузариум, пеницилл окрашивают древесину в розовый или желтый цвет, и т. д. Эти грибы развиваются в паренхимных клетках заболони за счет содержимого клеток и почти не разрушают клеточные стенки. Поэтому древесина на этой стадии, по существу, не изменяет свою структуру и физические свойства.
Вслед за этими грибами в древесину проникает мицелий базидиальных грибов. Его сопровождают так называемые грибы-спутники (пенициллы), которые могут питаться содержимым клеток и использовать промежуточные продукты, образующиеся при разрушении древесины (лигно-целлюлозного комплекса) базидиальными грибами. Грибы-спутники - активные кислотообразователи, содействующие подкислению среды. Это обеспечивает определенные экологические условия, при которых в древесине еще более пышно развивается мицелий основных дереворазрушителей - базидиальных и несовершенных грибов, способных разлагать клетчатку, т. е. видов родов триходерма, стахиботрис и других, а также некоторых сумчатых грибов. По мере разрушения клеточных стенок и исчерпания целлюлозы мицелий базидиальных грибов постепенно отмирает, частично разлагается с выделением аммиака и среда подщелачивается. Пенициллы исчезают, и появляются новые формы грибов, например виды родов гаплографиум (Haplographium), споротрихум (Sporotrichum) и других, при одновременном, более сильном развитии грибов, участвующих в предыдущих стадиях разложения (видов рода триходерма), способных к энергичному расщеплению клетчатки и белков, сильных аммонификаторов. На этой последней стадии разрушения колода теряет свою форму, покрывается мхами и другими растениями, а от древесины остается темная труха. Распад завершается появлением различных шляпочных грибов.
В зависимости от типа леса, породы дерева и других условий состав видов и родов грибов на разных стадиях разложения может несколько варьировать. Например, при разложении сосновой и еловой древесины развиваются разные виды пенициллов. Кроме того, в процессе разложения еловой древесины принимает участие большее количество видов этого рода и т. п.
Сходная картина, т. е. смена видов грибов, наблюдается при разрушении пиломатериалов и другой, так называемой деловой древесины.
Смена одних видов грибов другими имеет место и при разрушении лесных подстилок. Разным стадиям разложения подстилки соответствуют определенные комплексы грибов. В поверхностном слое присутствуют виды альтернария, кладоспорий, сферопсидные и другие грибы, представляющие собой несовершенные стадии сумчатых грибов. Они попадают в подстилку с растительными остатками (хвоей, листьями). Во втором слое, наиболее биогенном и активном, помимо основных разрушителей опада (сумчатых и базидиальных грибов), есть многочисленные несовершенные грибы - пенициллы, триходермы, аспергиллы и др.
Определенная флора несовершенных грибов характерна для разложения растительных остатков в полях или степях.
В последние годы установлено, что группа деревоокрашивающих грибов неоднородна по своему влиянию на древесину. Некоторые из них почти не изменяют ее свойства, другие могут сильно увеличивать водопоглощение, сопротивление ударному изгибу и даже существенно снижать прочность древесины. Есть среди них такие, которые способны переходить к разрушению клеточных стенок, по-видимому, после того, как будут использованы легкоусвояемые вещества содержимого клеток.
Такие изменения, как увеличение водопоглощения, образование отверстий в поперечных стенках клеток паренхимы, способствуют развитию в древесине мицелия базидиальных грибов - основных дереворазрушителей.
У представителей многих родов несовершенных грибов (Myrothecium verrucaria, Trichoderma viride, Trichoderma koningii, Stysanus stemonites, некоторые виды пенициллов) экспериментально установлена способность продуцировать целлюлозолитические ферменты. Это позволяет использовать их в целлюлозно-бумажной промышленности для получения определенных сортов бумаги и картона из древесных отходов.
Гифомицеты - паразиты растений (М. В. Горленко)
Среди гифомицетов известны многие паразиты, вызывающие заболевания растений (фитопаразиты). Некоторые из них могут быть причиной значительных потерь урожая возделываемых растений (пирикуляриоз риса, церкоспороз сахарной свеклы и др.). Чаще всего эти грибы образуют на листьях, стеблях или других органах некроз - пятнистость разного цвета. Некоторые гифомицеты бывают причиной сосудистых заболеваний, приводящих к увяданию растений (вилт хлопчатника и др.), гниению растительных тканей (корневая гниль злаков, фузариозная, или сухая, гниль клубней картофеля, серая гниль цветков, кочанов капусты).
Род Ботритис (Botrytis) (М. В. Горленко)
Грибы, относящиеся к роду ботритис, паразитируют на растениях, вызывая серую гниль различных частей их. При этом ботритис образует обильный серый налет, состоящий из мицелия и бесцветных или слабо-дымчатых одноклеточных конидий, размером примерно около 10 мкм. Отсюда и название болезни - серая гниль. Кроме конидий и мицелия, у многих видов есть также склероции, после перезимовки или прорастающие в мицелий, или у некоторых форм развивающие плодовые тела - апотеции с сумками и аскоспорами. Развитию плодовых тел предшествует половой процесс.
Центральное место в роде ботритис занимает многоядный паразит ботритис серый (В. cinerea) - возбудитель серой гнили многих растений. Особенно опасны серая гниль ягод винограда и земляники, кагатная гниль, поражающая сахарную свеклу во время хранения в кагатах, серая гниль кочанов капусты, стеблевая гниль гречихи, конопли и ряда других растений, гниль плодов тыквенных растений и некоторые другие. При высокой влажности этот гриб вызывает серую гниль цветков многих декоративных растений - пионов, гладиолусов и других, особенно при выращивании цветов в оранжереях.
Возбудитель серой гнили (табл. 59) - космополит, паразитирующий на разных растениях в большинстве районов земного шара. Но, вероятно, в составе популяции этого вида гриба существуют формы или расы, преимущественно поражающие какой-то определенный вид растения, а может быть, приуроченные к той или иной климатической зоне. Во всяком случае, одна из форм этого гриба - активный возбудитель гниения корней сахарной свеклы во время хранения - слабо поражает ягоды винограда. По-видимому, только некоторые формы возбудителя серой гнили образуют сумчатую стадию.
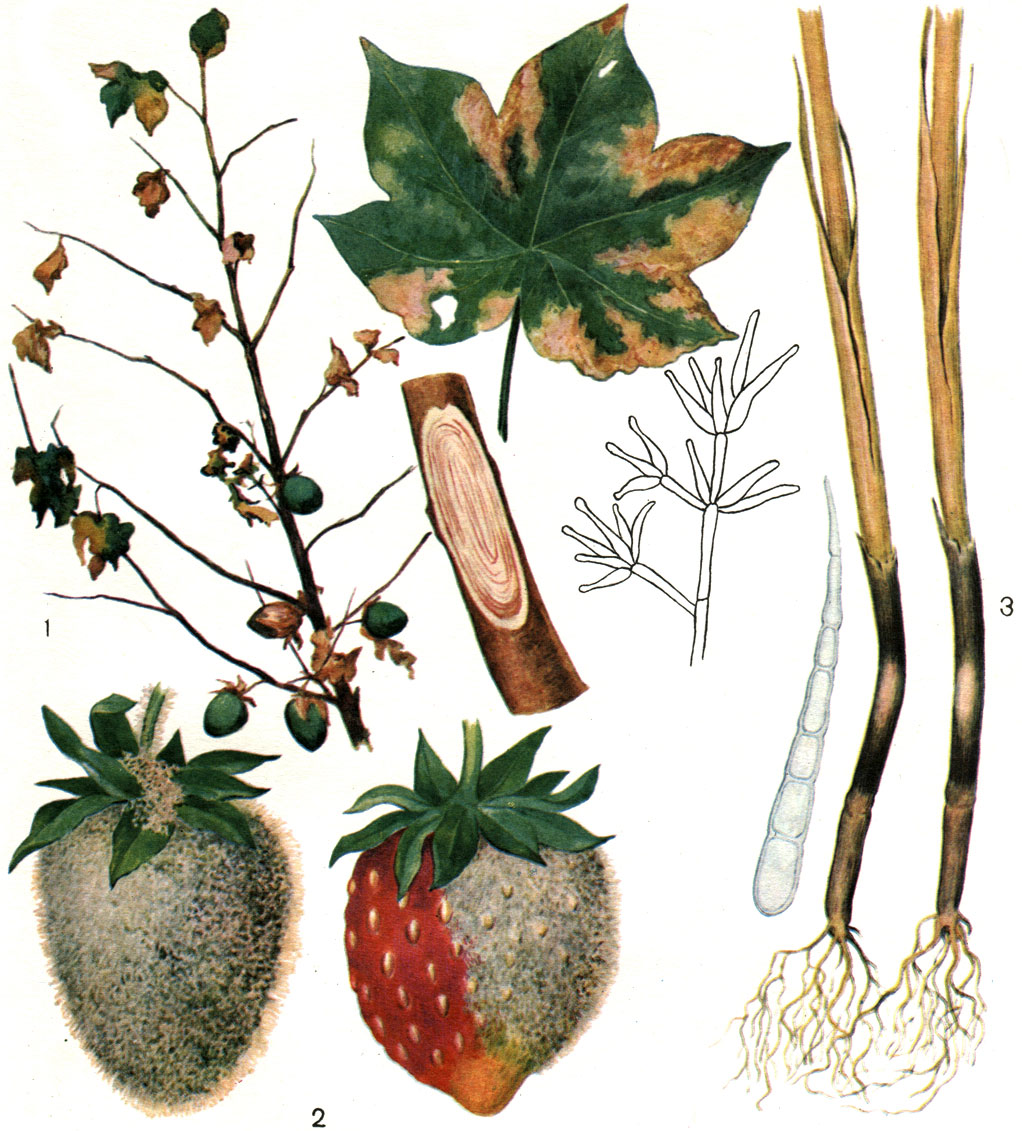
Таблица 59. Гифомицеты - паразиты растений: 1 - вертициллезный вилт хлопчатника (возбудитель - Verticillium dahliae), справа - спороношение гриба; 2 - серая гниль земляники (возбудитель - Botrytis cinerea); 3 - ломкость стеблей пшеницы (возбудитель - Cercosporella herpotrichoides), слева - конидия паразита
Серый ботритис живет в почве на растительных остатках в виде мицелия и склероциев. Так как гриб может развиваться на отмерших частях многих растений, то источников заражения им в природе всегда много. Развитие гриба с весны чаще всего начинается с разрастания мицелия и образования конидий на склероциях или на зараженных растительных остатках, на которых зимовал мицелий гриба. Конидии гриба инфицируют восприимчивые части растений.
У некоторых форм (или рас) склероции при прорастании образуют сумчатую стадию, и тогда растения заражаются аскоспорами. В нашей стране способность возбудителя серой гнили образовывать сумчатую стадию впервые обнаружена М. А. Кублицкой в Крыму у формы гриба, вызывающего серую гниль ягод и других частей растений виноградной лозы.
Особенности паразитизма у возбудителя серой гнили весьма своеобразны. Для того чтобы вызвать поражение живых тканей растений, он должен сначала поселиться хотя бы на небольшом участке отмершей ткани. После этого своими токсическими выделениями гриб отравляет соседние живые клетки, затем проникает туда и использует их для своего питания, отравляя тем временем следующую группу расположенных рядом живых клеток, постепенно захватывая все новые и новые живые ткани. Токсины паразита как бы идут впереди, подготавливая почву для его питания. Поэтому возбудителя серой гнили часто называют "паразитом теплого трупа", имея в виду, что он непосредственно для питания использует только что убитые им самим ткани.
Поражение винограда серой гнилью начинается уже с весны, с развития гриба на отмерших частях цветка (тычинки и т. п.), отмерших участках листовой пластинки, погибших усиках и т. п. Через некоторое время гриб переходит на живые ткани указанным выше способом.
Серая гниль широко распространена почти во всех районах виноградарства. Для борьбы с серой гнилью винограда применяют опрыскивание виноградников химикатами. В Молдавии выведены сорта винограда, устойчивые к серой гнили.
Во влажные годы от серой гнили гибнет много ягод земляники, иногда до 40-60%. У земляники, кроме ягод, серая гниль поражает бутоны, плодоножки и листья, на которых развиваются мокнущие пятна с характерным серым налетом. Ягоды могут заражаться из почвы, причем сначала заболевают те, которые ближе расположены к земле, а уже от них ягоды верхних ярусов. Гриб может развиваться сначала на остатках лепестков и тычинок, а уже оттуда через цветоложе грибница его проникает в ягоды. Земляника сильнее поражается серой гнилью в загущенных посадках и во время дождливой прохладной погоды.
Для борьбы с серой гнилью земляники нужно уничтожать остатки отмерших растений осенью или весной. Хорошие результаты дает комбинированное применение биологического препарата триходермина при внесении его в почву весной, а затем опрыскивание 1%-ным раствором препарата ТМТД в начале цветения.
Интересный гриб, поражающий цветки красного клевера, открыл в 1914 г. А. С. Бондарцев. Он назвал его ботритис цветолюбивый (Botrytis anthophila), а болезнь - цветочной плесенью. У больных растений пыльники покрываются серой плесенью, состоящей из бесцветных одноклеточных конидий возбудителя болезни. Сами растения внешне здоровы, заболевают лишь цветки. Споры гриба разносятся ветром или насекомыми. Попадая на рыльца здоровых цветков, конидии прорастают и грибница, продвигаясь по столбику, проникает в завязь, сохраняясь в семенах до будущего года. Зараженные семена щуплые, частично они гибнут, а из некоторых вырастают больные растения, внутри их тканей развивается грибница, проникающая в конечном итоге в генеративные органы и вызывающая поражение цветков. Через зараженные части цветков грибница может проникать в стебли и зимовать в их основании, а после отрастания растений развиваться внутри тканей, а достигнув точки роста, вызывать поражение новых цветков. Гриб приносит большой вред семеноводству клевера. Для борьбы с ним надо сортировать семена, освобождаясь от щуплых, а также протравливать их гранозаном.
Возбудитель цветочной плесени поражает, кроме культурных, также и дикие клевера, от которых возможно заражение культивируемых растений. Поэтому дикорастущие клевера следует скашивать до их цветения.
Весьма вредоносную болезнь лука - шейковую гниль - вызывает гриб ботритис луковый (Botrytis allii). Он поражает луковицы, а иногда также цветоносные побеги и соцветия семенников. У луковиц обычно загнивает шейка или донце, а затем и вся луковица. Пораженные ткани покрываются серой плесенью, состоящей из массы мицелия и одноклеточных бесцветных конидий гриба. Позднее на больных луковицах развиваются черные склероции, часто сливающиеся в сплошную черную корочку. Пораженные луковицы гибнут. Лук заражается шейковой гнилью еще в поле. Однако основной вред болезнь приносит во время хранения лука. Источником заражения здоровых луковиц служит главным образом посадочный материал. Это связано с тем, что среди высаживаемых луковиц всегда попадается хотя бы небольшое количество зараженных шейковой гнилью.
Попав непосредственно в почву, возбудитель шейковой гнили там довольно быстро гибнет, так как очень чувствителен к воздействию почвенных микроорганизмов. Этот гриб зимует в зараженных луковицах. Затем уже в поле происходит перезаражение здоровых растений. Гриб сначала развивается за счет отмерших или отмирающих частей растений и, проникая в шейку, не вызывает первоначально ее загнивания. Болезнь начинает проявляться лишь через месяц-полтора после закладки лука на хранение. На уже зараженных еще в поле луковицах болезнь быстро прогрессирует, а от развивающихся на них спор заражаются и здоровые луковицы.
Заражение лука шейковой гнилью в поле скорее всего происходит при высокой влажности почвы, во влажную погоду и на тяжелых суглинистых почвах. Быстрое подсыхание листьев препятствует проникновению гриба в шейку луковицы. Поэтому для борьбы с шейковой гнилью надо применять мероприятия, ускоряющие вызревание лука, проводить просушку лука после уборки до подсыхания листьев, подсохшие листья обрезать. Важно также соблюдать режим хранения. Температура в хранилищах должна быть 0-3° С, влажность - 75-80%.
Грибы из рода ботритис приносят большой вред цветоводству, особенно луковичным растениям. На луковицах и других частях растений тюльпанов паразитирует гриб ботритис тюльпанный (В. tulipae), на нарциссах - ботритис нарциссовый (В. narcissicola). Особенно опасно загнивание луковиц этих растений во время хранения и транспортировки. Поражаются имп и вегетативные органы. Бобам сильно вредит ботритис бобовый (В. fabae), поражающий вегетативные органы этих растений
Для грибов рода вертицилл характерны следующие признаки. Конидиеносцы прямостоящие, с перегородками, мутовчато разветвленные; на них расположены фиалиды, часто бутыльчатой формы и на концах заостренные. На фиалидах видны одиночные или скопляющиеся в головки конидии, иногда склеенные слизью, быстро распадающиеся. Форма их разнообразна: шаровидные, яйцевидные, эллипсоидальные, веретеновидные. Они либо бесцветны, либо светло окрашены. Стерильные гифы ползучие, бесцветные или окрашенные. У этих грибов есть хламидоспоры, геммы, склероции, микросклероции и дауэрмицелий. При помощи их грибы переносят неблагоприятные условия и сохраняют способность к дальнейшему самовоспроизведению. В числе видоизменений мицелия вертициллов некоторые авторы называют оидии, или оидиоподобные образования (рис. 233).
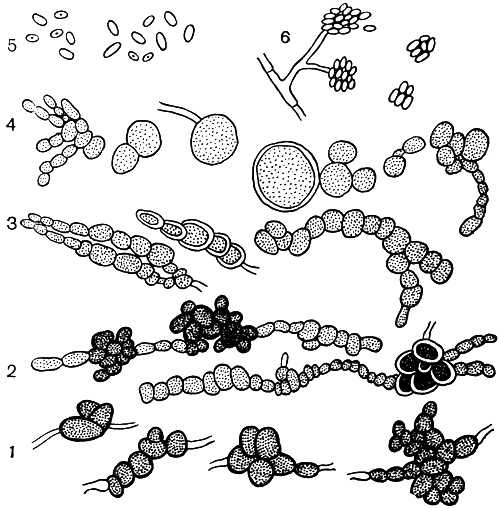
Рис. 233. Типы спороношения грибов из рода вертицилл (Verticillium): 1 - микросклероции; 2 - дауэрмицелий с микросклероциями; 3 - дауэрмицелий; 4 - оидиоподобные образования; 5 - фиалоспоры; 6 - конидиеносец и конидии
Хламидоспоры, геммы, склероции часто встречаются у грибов разных систематических групп; образование же дауэрмицелия и микросклероциев - явление довольно редкое. Дауэрмицелием называют темные толстостенные гифы, с возрастом уплотняющиеся. Дауэрмицелий может снова образовывать стерильные бесцветные или окрашенные гифы, либо конидиеносцы с конидиями. Микросклероции - это мелкие или крупные, иногда удлиненные, плотные ячеистые тела различной формы. Однако форма, плотность, количество и размеры их очень изменчивы и зависят от многих причин. В микросклероциях имеются два типа клеток: толстостенные темные и тонкостенные бесцветные. Функция толстостенных - защита и накопление питательных веществ для тонкостенных, а функция тонкостенных - сохранение вида (выживание его) при неблагоприятных условиях. Вероятно, с этим связано и их цитоплазматическое отличие: толстостенные клетки состоят из различных цитоплазменных включений, вакуолей и не имеют морфологически дифференцированного ядра, а тонкостенные клетки содержат одно или несколько ядер.
В цикле развития грибов рода вертицилла чередуются вегетативный рост мицелия, бесполое размножение при помощи конидий и покоящиеся стадии. При наступлении благоприятных условий покоящиеся структуры (геммы, хламидоспоры, склероции, микросклероции) прорастают мицелием, на котором образуются конидиеносцы с конидиями.
Среди грибов рода вертицилл есть паразиты растений и сапрофиты. Большинство из них приурочено к растительным субстратам (живым или гниющим). Поражая здоровые растения, некоторые виды вертициллов вызывают их увядание (вилт) или усыхание. Основной симптом этого заболевания - потеря тургора листьями, появление темной окраски проводящих сосудов и гибель либо всего растения, либо его отдельных частей. Подобное явление может быть вызвано другими грибами, а также бактериями или недостатком влаги, но при вертициллезном поражении из больных тканей всегда выделяются колонии тех или иных вертициллов.
Гниение ягод винограда вызывает V. ampelinum, а желудей во время вегетации и хранения - V. candelabrum.
Несколько видов (до 10) этого рода являются микофильными грибами, т. е. паразитируют на других грибах. Они могут поселяться на сыроежках, свинушках, лисичках, рогатиках, деревбразрушающих грибах, представите-лях рода болет (Boletus) и других родов.
На шляпках грибов появляется беловатый или розоватый войлочный мицелий, и они загнивают или засыхают, становясь непригодными к употреблению. Например, V. psalliotae вызывает гибель шляпочного гриба Rhopalomyces elegans при его промышленном разведении. Даже в споровместилищах слизевиков встречается V. niveo-stratosum. Этот же гриб находили и на гниющих пнях.
Многие вертициллы поселяются на коре деревьев, на сплавной и деловой древесине. Эти грибы обитают в почве, в воде стоячих и проточных водоемов. Гриб V. aphidis известен как паразит тлей.
Экологический спектр некоторых видов достаточно широк: часто грибы одного вида могут быть обнаружены на самых различных субстратах. Например, V. candelabrum найден на древесине гниющего дуба, на засохших листьях различных пород деревьев и в болотной воде, а V. glaucum - на сплавной и гниющей древесине, на коре деревьев, на сене, в почве. Все это свидетельствует о большой пластичности видов этого рода.
Из всего многообразия представителей рода Verticillium наибольшее практическое значение имеют фитопатогенные виды, особенно паразиты, вызывающие увядание растений. Они встречаются как в северных районах нашей страны, так и в южных (Молдавия, Закавказье, Средняя Азия). Эти грибы поражают важнейшие сельскохозяйственные культуры, например хлопчатник, подсолнечник, плодовые, цитрусовые и т. д.
Увядание (вилт) хлопчатника распространено во всех районах хлопкосеяния. Чаще всего первые признаки заболевания появляются в фазе бутонизации, сначала на нижних листьях, как пожелтение отдельных участков листовой пластинки между жилками. Тогда заболевшие листья поникают и опадают. Постепенно увядание охватывает все листья. Растения приостанавливаются в росте, коробочки на них подсыхают и преждевременно раскрываются. Иногда возможно и быстрое увядание растений, когда они погибают без видимых причин заболевания. Как при хроническом течении болезни, так и при молниеносном увядании в зараженных растениях обнаруживают потемнение сосудов (табл. 59).
Побурение и потемнение сосудов проводящей системы больных растений - отличительный признак вертициллезного увядания. В пораженных сосудах обнаруживается мицелий гриба, скопление камеди - гуммиобразного вещества, закупоривающего сосуды. Иногда наблюдается образование тилл (пузыревидных выростов боковых стенок сосудов), сужающих просветы их. Подобное поражение водопроводящей сосудистой системы растения-хозяина позволило первоначально предположить, что причина увядания кроется в закупорке сосудов мицелием гриба, что препятствует продвижению воды и питательных веществ. Впоследствии эта гипотеза уступила место мнению о токсической природе вилта растений. Продуцируемые возбудителем токсины нарушают физиологические процессы в растении, влияя на различные стороны его обмена веществ, что приводит к гибели растения. Относительно природы токсинов существуют различные предположения.
Широко распространено вертициллезное увядание плодовых культур. В пределах СССР на них зарегистрировано 13 паразитных и сапрофитных видов вертицилла. Кроме Verticillium albo-artrum и V. dahliae, паразитами плодовых деревьев являются V. cornicolor, V. ibericum, V. nigrescens и др.
Ареал заболевания охватывает территорию между 30 и 35° северной и южной широты. Оно может встречаться в умеренном (США, юг Канады, Франция, Молдавская ССР), субтропическом (Калифорния, Югославия, Грузинская ССР) и тропическом поясах (некоторые районы Австралии). Наиболее часто болезнь охватывает насаждения косточковых плодовых деревьев (персик, абрикос, миндаль, слива, табл. 63) и значительно реже встречается на семечковых породах (яблоня, груша). Обычно вилт проявляется в середине лета, когда у пораженных деревьев начинают желтеть и опадать листья с плодовых веточек у основания скелетных ветвей. Впоследствии хлороз распространяется на молодой прирост и на всю крону. Внешним признакам вертициллезного усыхания сопутствует некроз древесины. Он заметен не только на поперечных, но и на продольных срезах, в виде прерывистых или сплошных темных широких полос.

Таблица 63. Вертициллезное увядание сливы: справа - больное дерево; слева вверху - продольный, внизу - поперечный срез больной древесины
Вертициллезным увяданием могут поражаться плодовые деревья в любом возрасте, но наиболее восприимчивы молодые посадки (от двух до пяти лет). Заболевание бывает чаще распространено очагами.
Основной возбудитель увядания растений - V. dahliae - поражает свыше 350 представителей из многих семейств. Наиболее часто этим грибом бывают поражены картофель, томаты, баклажаны, различные тыквенные, астры, львиный зев, дурнишник, сурепка, вьюнок полевой, клен, пальма, шелковица и т. д.
Гриб внедряется в растение чаще через корни, но мицелий обнаруживается в стеблях, черешках, листьях, а у некоторых - в плодах и семенах. Остатки зараженных растений, попадая в почву, становятся источниками накопления, распространения и сохранения инфекции на довольно продолжительное время.
Гриб сохраняется на растительных остатках в виде микросклероциев, обильно образующихся при попадании в почву. Имеются сведения о сохранении гриба в форме микросклероциев на отмерших растительных остатках от 5-6 до 14-15 лет, а в виде спор и мицелия - в течение нескольких дней или недель (не более 3-4месяцев). На выживание в почве микросклероциев оказывает влияние комплекс условий, создающихся в ней. Ведущую роль при этом играют устанавливающиеся взаимоотношения между сапрофитной и патогенной микофлорой. Жизнедеятельность почвенных микроорганизмов тормозит прорастание инфекционных зачатков (в нашем случае - микросклероциев). Это предохраняет их от преждевременного прорастания и гибели. До тех пор, пока микросклероции находятся в состоянии покоя, они неуязвимы для окружающей микрофлоры.
Состояние покоя (фунгистазиса) может быть преодолено корневыми выделениями растений, стимулирующими жизнедеятельность инфекционных зачатков. Действие корневых выделений малоспецифично, т. е. прорастание микросклероциев может быть вызвано корневыми выделениями многих растений, в том числе и непоражаемых грибом. Как только микросклероции прорастают, мицелий, развивающийся из них, подвергается воздействию почвенной микрофлоры - лизирующих бактерий, грибов-антагонистов и т. д. Они либо убивают разрастающийся мицелий, либо задерживают его развитие. Если прорастание вызвано корневыми выделениями восприимчивого растения, то грибница может "уйти" от губительного действия сапрофитной микрофлоры, внедрившись через корни в растение и вызвать его гибель или угнетение.
В связи с этими особенностями поведения возбудителя вертициллезного увядания в почве наиболее эффективной мерой борьбы с ним является построение правильных севооборотов. В них должно быть предусмотрено возвращение одноименных культур на прежнее место через несколько лет и подобраны непоражаемые предшественники. В частности, хорошее оздоровляющее действие в борьбе с вилтом хлопчатника оказывают хлопково-люцерновые севообороты или введение в севообороты других иммунных, например злаковых, культур с получением двух урожаев в год. В плодовых садах, в междурядьях, также рекомендуется высаживать невосприимчивые растения и ни в коем случае нельзя возделывать пасленовые, подсолнечник и землянику, так как они неустойчивы к возбудителю. Под новые плодовые насаждения следует отводить те участки, где длительное время не возделывали овощные культуры, многие из которых восприимчивы к возбудителю вилта. Более подробно рекомендации по борьбе с вертициллезным увяданием растений описаны в специальных руководствах.
Несколько видов рода вертицилла известны на корнях сахарной свеклы. При повышенной температуре почвы одни из них вызывают загнивание ткани корнеплодов в почве на глубину 1,5-3,0 см. Свое развитие они продолжают и при хранении свеклы в кагатах, где к ним присоединяются другие виды. Их появление хорошо заметно в виде кирпично-красного налета на срезанных головках корней. При хранении свеклы виды вертициллов чаще выступают как сапрофиты, в качестве спутников кагатной гнили, развиваясь на уже загнивших корнеплодах и в симбиозе с другими микроорганизмами.
К роду церкоспорелла относят гифомицеты, образующие удлиненно-булавовидные или цилиндрические бесцветные конидии с многочисленными поперечными перегородками. Они образуются на простых или слабоветвящихся конидиеносцах, выходящих пучками из устьиц на нижней стороне пораженных грибом листьев растений. Многочисленные виды этого рода паразитируют на листьях растений, вызывая у них образование пятен различной формы, обычно желтоватых или буроватых, окруженных каймой.
Один из наиболее важных видов этого рода - церкоспорелла герпотрихоидес (Cercosporella herpotrichoides). Этот гриб вызывает опасное заболевание хлебных злаков - эллипсовидную, или глазковую, пятнистость, прикорневую гниль или ломкость стеблей. Оно широко распространено в ряде стран Европы (Англия, Франция, Голландия, Бельгия, Австрия, ГДР, ФРГ, Польша, Болгария), в Северной Америке и Австралии, преимущественно в более влажных районах. Гриб поражает пшеницу, рожь, ячмень, многие кормовые и дикорастущие злаки. В отдельные годы при сильном развитии болезнь вызывает большие потери урожая (до 30-40%). В нашей стране эта болезнь встречается в некоторых западных районах европейской части и на Северном Кавказе.
Церкоспорелла герпотрихоидес сильнее всего поражает озимую пшеницу. В нижней части пораженных стеблей, на первом, а реже и втором междоузлии, образуются светлые овальные пятна длиной до 1-2 см, окруженные темной каймой и особенно хорошо заметные на листовом влагалище (табл. 59). Иногда они опоясывают все основание стебля. На продольных разрезах пораженных стеблей хорошо заметен серый мицелий гриба, заполняющий их полость. Зараженные стебли искривляются и переламываются. В результате этого происходит полегание растений. При сильном поражении растений уменьшаются размер колосьев и масса зерна, часто наблюдается белоколосость.
Паразит зимует на зараженных пожнивных остатках, а также дикорастущих злаках (мятлики, плевел и др.), которые служат источником инфекции весной. В период вегетации заражение растений вызывают конидии, переносимые с больных растений на здоровые воздушными течениями.
У представителей рода пирикулярия бесцветные или слегка коричневатые конидии с двумя или большим числом перегородок, образуются на вершине простых или слабо разветвленных конидиеносцев группами (рис. 234). Все представители этого рода паразитируют на цветковых растениях, вызывая у них пятнистость листьев.
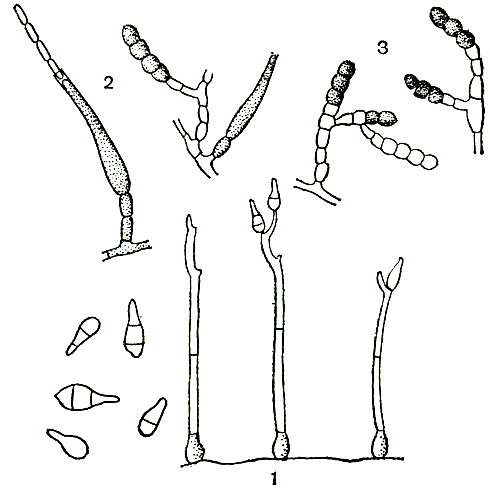
Рис. 234. Гифомицеты: 1 - пирикулярия рисовая (Piricularia oryzae); 2, 3 - тиелавиопсис корневой (Thielaviopsis basicola)
Пирикуляриоз риса, вызываемый пирикулярией рисовой (Piricularia oryzae),- одно из наиболее распространенных и вредоносных заболеваний этой культуры. Впервые оно было описано в Японии в 1704 г., а его возбудитель - в 1896 г. Сейчас пирикуляриоз известен во всех районах возделывания риса.
Заражение растений может происходить в разных фазах развития, но более восприимчивы к пирикуляриозу молодые растения. При заражении растений в ранние фазы развития (до выхода в трубку) развивается листовая форма пирикуляриоза. На листьях появляются светлосерые пятна, при сильном поражении растения засыхают до образования метелок. При заражении растений перед выметыванием метелок гриб поражает преимущественно узлы стебля, а начиная с этой фазы - все части метелок и колосковые чешуи. С нижней стороны пятен развивается конидиальное спороношение гриба. Его грушевидные трехклеточные конидии с придатком образуются на симподиально ветвящихся конидиеносцах. Сначала образуется конидия на вершине конидиеносца, затем рядом - новая точка роста, развивается веточка конидиеносца, а на ней - следующая конидия; этот процесс повторяется до 7-9 раз. Образование последовательных конидий происходит с интервалами около часа.
Наибольший вред приносит пирикуляриоз при его развитии в период колошения и цветения риса, особенно если поражается основание главной оси метелки. Метелки в этом случае часто высыхают до образования зерна или в них образуются щуплые, недоразвитые зерновки. Даже при слабом поражении пирикуляриозом потери урожая достигают 20-30%, а в периоды эпифитотий они значительно превышают эти цифры.
Возбудитель пирикуляриоза образует фитотоксины - пирикулярин и α-пиколиновую кислоту, вызывающие у растений типичные симптомы заболевания. Устойчивые к пирикуляриозу сорта риса устойчивы и к действию этих токсинов.
Пирикулярия рисовая заражает многие злаковые растения (например, костер, просо, тимофеевку, пырей, ежу, душистый колосок). Однако в пределах этого вида наблюдается дифференциация на биологические расы, различающиеся по патогенности для различных сортов риса. Известны 32 расы этого гриба, соотношение которых в популяциях паразита ежегодно меняется.
Сохранение жизнеспособности паразита в растительных остатках в сильной степени зависит от условий перезимовки. На погруженных в почву остатках он погибает к весне. Если зараженная солома долго находится под снегом или под продолжительными дождями, пирикулярия рисовая также быстро погибает. В сухой соломе ее мицелий сохраняет жизнеспособность в течение 2-4 лет, особенно в узлах стеблей. После дождей или росы на нем появляются конидии, заражающие молодые растения. Инфекция хорошо сохраняется и на семенах. Источником заражения могут служить и дикорастущие злаки, пораженные пирикуляриозом. В некоторых странах резерватором пирикулярии служат сорняки на рисовых полях (США, Бразилия, Индия). В других странах (например, в Японии) перезимовку паразита на живых растениях не наблюдают.
В связи с большими потерями урожая риса от пирикуляриоза во всех странах с развитым рисоводством разрабатываются мероприятия по борьбе с этой болезнью. Получены устойчивые к ней сорта риса, но многие из них после внедрения в производство быстро теряют свою устойчивость вследствие появления новых рас паразита. Для борьбы с пирикуляриозом рис обрабатывают антибиотиками (бластицидином, касугамицином и др.).
Другой вид этого рода - пирикулярия серая (P. grisea) - вызывает серую пятнистость проса, чумизы и щетинника. В 1972 г. в культуре этого гриба была получена сумчатая стадия цератосфера серая (Ceratosphaera grisea), относящаяся к порядку диапортовых. У пирикулярии рисовой сумчатая стадия неизвестна, хотя некоторые микологи предполагают, что она связана с родом микосферелла (Mycosphaerella) из локулоаскомицетов.
Для грибов рода кладоспорий характерно конидиальное спороношение в виде деревца, ветви которого состоят из спор (рис. 235). Ствол "деревца" образован буроватым или бледно-оливковым конидиеносцем, обычно прямостоячим, септированным, неветвящимся или редко с 1-2 боковыми веточками. "Крона" состоит из коротких разветвленных цепочек конидий, которые создают впечатление боковых ветвей. Наличие таких боковых цепочек спор указывает на то, что вершина конидиеносца продолжала свой рост после образования первых конидий. Непосредственно от конидиеносца отходят длинные цилиндрические конидии. Их называют базальными конидиями или метаконидиями. Метаконидии дают начало более коротким продолговато-эллиптическим или цилиндрическим спорам, которые, в свою очередь, отпочковывают одноклетные конидии яйцевидной, овальной или округлой формы. Таким образом, конидии на одном конидиеносце отличаются по форме и величине. Оболочка конидий бывает гладкой или шиповатой. Молодые конидии всегда гладкие, бесцветные, одноклетные. При их созревании у многих видов оболочка становится шиповатой и появляются поперечные перегородки. Окраска варьирует от бледно-оливковой до бурой. Конидии прорастают через 5-6 ч одной или двумя, реже тремя ростковыми трубками. Отдельный конидиеносец несет от 100 до 300 конидий в зависимости от вида гриба. Грибы, паразитирующие на растениях, имеют более крупные конидии, и количество их на конидиеносце значительно меньше, чем у сапрофитных видов.
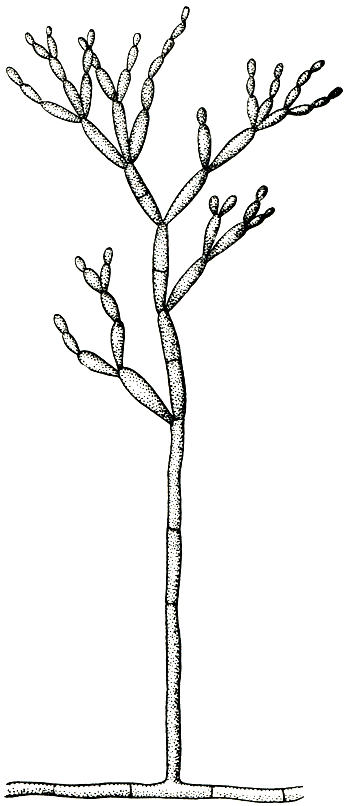
Рис. 235. Кладоспорий (Cladosporium)
Сумчатое спороношение обнаружено только у двух видов кладоспория. У них образуются плодовые тела типа перитеция с короткой шейкой (роды микосферелла и аморфотека). Сумки в перитеции содержат по восьми аскоспор.
Мицелий у кладоспория, как и споры, буровато-оливковый. Окраска в значительной мере определяется меланиновыми пигментами, образующимися в клетке в результате ферментативного окисления тирозина или полифенолов. Эти пигменты определяют устойчивость мицелия и спор к облучению. Поэтому грибы темного цвета хорошо развиваются на поверхности растений и других субстратов не только в зоне умеренного климата, но в пустынях и полупустынях.
По широте распространения кладоспорий можно отнести к числу наиболее часто встречающихся родов несовершенных грибов. Его представители обнаружены на самых разнообразных субстратах растительного и животного происхождения как в качестве сапрофитов, так и практически важных паразитов растений. Опасными патогенами растений являются, например, кладоспорий бурый (С. fulvum, табл. 61) - возбудитель бурой пятнистости томатов; кладоспорий огуречный (С. cucumerinum) - возбудитель оливковой плесени огурцов.
Наиболее многочисленны и широко представлены в этом роде сапрофитные виды - оливково-зеленые плесени. Они часто встречаются на отмирающих на корню растениях и на всевозможных растительных остатках, играя в одних случаях положительную роль, в других - отрицательную. Кладоспорий травяной (С. herbarum) и другие сапрофитные виды часто развиваются (особенно после влажных сезонов) на зерновках злаков и вызывают почернение зерна, а попав в хранилище,- его порчу. Если злаки зимуют под снегом (например, пшеница, рожь, просо), то мицелий кладоспория проникает в зерно и делает его токсичным для человека и животных. Многие грибы рода появляются сначала на отмирающих растениях, а затем, попав в хранилища, являются причиной порчи сена даже в условиях слабо повышенной влажности.
Кладоспорий заселяет не только отмерший растительный материал. Он весьма обычен на здоровых растениях как постоянный компонент эпифитной микробной флоры зрелых листьев растений. Установлено, что кладоспорий травяной, кладоспорий крупноспоровый (С. macrocarpum) и другие встречаются эпифитно на листьях разных злаков, древесных пород, овощных и ягодных культур, на листьях сахарного тростника и многих других растений, находясь там в активном состоянии, вегетируя и размножаясь.
Кладоспорий обитает в почве преимущественно на растительных остатках. Многие его виды обнаружены в торфах и в ризосфере растений. Кладоспорий травяной и другие грибы этого рода изобилуют в лесной подстилке, участвуя в ее разложении.
Споры кладоспория найдены в осадочных породах на глубине 18-1127 м в океане, в янтаре и на древесине в третичных отложениях, что свидетельствует о значительной древности этого рода.
В связи с широким распространением видов кладоспория на растениях и в почве большое количество его спор находится в воздухе. Особенно их много летом, в период вегетации растений (бывает более 40% всех обнаруженных в воздухе грибных спор). А в тропических массах воздуха количество спор достигает 82,3%.
Ввиду наличия большого количества спор кладоспория в воздухе не удивительна частая встречаемость видов этого рода на самых разнообразных субстратах, где эти грибы могут получать хотя бы незначительное количество питательных веществ. Они развиваются на жидком топливе, смазочных материалах, полихлорвиниловых покрытиях промышленных изделий в странах с тропическим климатом, на картинах, бумаге, древесине, на спороношениях некоторых базидиальных и сумчатых грибов. Они хорошо растут при пониженных температурах и часто встречаются на мясных продуктах, сливочном масле, упакованных овощах и фруктах при холодном хранении. При благоприятных условиях кладоспорий быстро размножается, обильно заселяя субстрат, и приносит значительный вред.
Описано около 300 видов кладоспория. Из них наибольший интерес представляют следующие виды.
Кладоспорий бурый (С. fulvum) вызывает распространенную в теплицах болезнь томатов - бурую пятнистость, или кладоспориоз. При тепличной культуре томатов бурая пятнистость приводит к значительным потерям урожая. В открытом грунте болезнь менее вредоносна и встречается в местах с влажным климатом. Возбудитель поражает главным образом листья, реже плоды и другие части растения. На пораженных листах появляются желтые с размытыми краями хлоротичные пятна. Этим пятнам с нижней стороны соответствует бархатистый буровато-оливковый налет спороношения гриба. Высокая влажность в теплице способствует развитию болезни. Обычно через 1-1,5 месяца после появления первых пятен все листья растений бывают сильно поражены, и это замедляет образование завязей и рост плодов. Снижение урожая зависит от сроков появления болезни и степени ее развития. Кроме того, сильная заспоренность теплиц может вызывать аллергические заболевания у работающего в теплице персонала. Наиболее действенная мера борьбы с этой болезнью - введение в культуру устойчивых сортов. В настоящее время известно 5 генов устойчивости к возбудителю бурой пятниогости, полученных от диких видов томатов. Сорта, обладающие одним или несколькими такими генами, обеспечивают устойчивость к определенным расам возбудителя, которых известно в настоящее время более 10.
Кладоспорий огуречный (С. cucumerinum) вызывает оливковую или бурую пятнистость огурцов. Больше всего он поражает плоды. На них образуются углубленные язвы неправильной формы, покрытые оливковым налетом спороношения гриба. При раннем заражении плоды искривляются и прекращают рост. На стеблях и черешках листьев пятна углубленные, на листьях - светло-бурые с оливковым налетом. Это заболевание представляет большую опасность при культуре огурцов в закрытом грунте, особенно при пониженной температуре и высокой влажности. Правильный режим в теплицах и выращивание устойчивых сортов позволяют бороться с этой болезнью.
Кладоспорий травяной (С. herbarum) - наиболее часто встречающийся вид, который развивается на самых разнообразных органических субстратах. Растет он плотными бархатистыми дерновинками оливкового, черного или зеленоватого цвета. Гриб разрушает сырье, содержащее целлюлозу и пектин, изменяет окраску бумаги и поверхностных слоев древесины. Это один из главных компонентов микрофлоры при росяной мочке льна.
Кладоспорий смоляной (С. resinae) морфологически несколько отличается от большинства других видов. Окраска его колоний песочного цвета, так как меланиновых пигментов в клетках не образуется. Существует гриб в виде двух разных конидиальных форм (f. avellaneum и f. resinae). Наряду с конидиальным спороношением он в культуре образует перитеции (Amorphotheca resinae). Гриб встречается в почве и является естественным компонентом воздушной флоры. Он отличается способностью легко усваивать углерод из углеводородов, поэтому хорошо растет на керосине, креозоте, дизельных топливах и различных смазках, вызывая разрушение этих веществ, отсюда известен под названием "керосинового" гриба.
К роду тиелавиопсис относят грибы, образующие в цикле развития две стадии: эндогенные фиалоконидии, обычно цилиндрические, бесцветные, развивающиеся цепочками в фиалидах, имеющих форму лампового стекла, и темноцветные, бочонковидные или цилиндрические хламидоспоры с толстыми стенками, развивающиеся цепочками на концах гиф (рис. 234). Представители этого рода обитают как паразиты или сапрофиты на корнях растений. Некоторые из них образуют сумчатые стадии, относящиеся к порядкам эуроциевых и микроасковых.
Наиболее распространенный вид этого рода - тиелавиопсис корневой (Thielaviopsis basicola), вызывающий черную корневую гниль различных растений и встречающийся во всех районах земного шара. Этот вид поражает более 100 видов растений, особенно страдают от него табак, хлопчатник, многие бобовые и другие растения. Злаки и ряд других растений устойчивы к нему. Строгая специализация у этого вида отсутствует. Изоляты, полученные с табака, могут, например, заражать хлопчатник.
В цикле развития тиелавиопсиса корневого развиваются фиалоспоры и хламидоспоры. Его конидии имеют тонкую стенку и быстро гибнут. Они служат только для размножения гриба в период вегетации. Основным источником инфекции являются хламидоспоры. Они образуются на концах гиф четковидными цепочками по 3-6, имеют темно-бурую окраску и толстую бородавчатую клеточную стенку (рис. 234). Хламидоспоры этого гриба представляют структуры, в процессе эволюции хорошо приспособ-ленные к перенесению неблагоприятных условий и к прорастанию в присутствии растения-хозяина. В лаборатории они быстро прорастают без периода покоя и воздействия низких температур. Однако в почве в отсутствие растения-хозяина они находятся в состоянии покоя под влиянием почвенного фунгистазиса. (Фунгистазис почвы - широко распространенное явление, состоящее, в частности, в подавлении прорастания спор, склероциев и других структур фитопатогенных грибов и связанное с выделениями почвенных микроорганизмов.)
Прорастание хламидоспор происходит лишь под влиянием корневых выделений растений. Однако прорастание хламидоспор тиелавиопсиса корневого стимулируют корневые выделения не только восприимчивых к заражению растений, но и устойчивых, например злаков. Под их воздействием хламидоспоры прорастают и быстро погибают от влияния почвенных антагонистов. Высевая пшеницу или другие злаки, можно таким образом очистить почву от заражения этим грибом.
Предполагают, что этот вид имеет сумчатую стадию тиелавия корневая (Thielavia basicola) из порядка эуроциевых. Но сумчатая стадия у этого гриба встречается, по-видимому, очень редко.
Черная корневая гниль табака, вызываемая грибом тиелавиопсисом корневым, распространена во всех районах мирового табаководства. Она поражает растения на всех стадиях развития, но чаше встречается на рассаде. При посеве табака в зараженную хламидоспорами гриба парниковую смесь они прорастают и заражают корни рассады. Пораженные корни сначала буреют, а затем чернеют. Листья больных растений увядают и желтеют, и растения часто гибнут. На пораженных корнях развиваются фиалоспоры и хламидоспоры паразита. При более слабом поражении растений в области корневой шейки образуются новые корешки и рассада внешне выздоравливает. Однако при высадке в поле такие растения могут быть источником инфекции и заразить здоровую рассаду. Кроме того, больные растения значительно отстают в росте и дают меньший урожай, и притом низкого качества.
При развитии черной корневой гнили в поле на корнях пораженных растений образуются темно-бурые пятна и корни отмирают. Сильное развитие болезни часто наступает через 2-3 недели после высадки рассады в поле. Причиной заболевания растений в поле может быть заражение почвы хламидоспорами паразита или высадка больной рассады. В почве хламидоспоры быстро гибнут при ее сильном высушивании и нагревании, однако во влажной почве их жизнеспособность сохраняется более двух лет. Поэтому черная корневая гниль табака в полевых условиях распространена только в тех районах, где не происходит продолжительного высушивания почвы. В парниках хламидоспоры паразита накапливаются в парниковой смеси, сохраняя жизнеспособность в течение нескольких лет. Особенно сильно поражается рассада при загущенном посеве и обильном поливе.
На интенсивность развития заболевания влияют температура и кислотность почвы. Растения поражаются сильнее при температуре 21-23° С, хотя в культуре возбудитель хорошо растет и при более высокой температуре; однако в последнем случае растение-хозяин хорошо развивается и болезнь вызывает меньший ущерб.
Относительно недавно черная корневая гниль начала распространяться на хлопчатнике. Эта болезнь поражает всходы и взрослые растения. При поражении всходов бывает и их массовая гибель. Листья больных растений теряют тургор и увядают, но сохраняя зеленую, окраску и не опадая. Корневая шейка утолщается, а на срезе нижней части стебля и корня хорошо видно изменение окраски ткани до малиновой или малиново-бурой. При сильном поражении происходит мацерация коры. У взрослых растений заболевание часто проявляется только к концу августа. Нередко кусты внезапно увядают и засыхают. Растения, пораженные черной корневой гнилью, плохо растут; коробочки на них не образуются, или их количество уменьшается; ухудшается также качество волокна.
Развитию болезни способствует температура почвы 17-24 °С и ее влажность около 60% (А. П. Xитрова). Кроме того, болезнь сильнее проявляется также при слишком раннем посеве хлопчатника.
Род церкоспора относят к семейству демациевых, хотя у многих его представителей мицелий и споры светлые. Для этого рода характерны бесцветные или окрашенные конидии, цилиндрической, обратнобулавовидной, веретеновидной или червеобразной формы с несколькими поперечными перегородками, вверху обычно утончающиеся. Они образуются на вершине простых, прямых или извилистых конидиеносцев, обычно окрашенных, одиночных или соединенных в коремии или ложа. В роде церкоспора насчитывают до 900 видов. Все представители этого рода - паразиты растений, вызывающие у них пятнистости на листьях, черешках, стеблях и плодах. Настоящие сапрофиты в этом роде неизвестны, хотя некоторые виды его встречаются как вторичные паразиты на растениях, ослабленных другими патогенами.
Листовая пятнистость, вызываемая церкоспорой, может быть слабо выраженным изменением окраски, но чаще это хорошо ограниченные пятна. При массовом развитии пятен листья погибают, а в некоторых случаях листья опадают полностью. Ткань пятен часто засыхает и выпадает из здоровой ткани листа, и листья становятся дырчатыми (рис. 236).
Рис. 236. Церкоспора свекольная (Cercospora beticola): 1 - пораженный лист свеклы; 2 - конидиеносцы с конидиями
Грибы этого рода поражают многочисленные травянистые и древесные растения, вызывая в некоторых случаях экономически важные заболевания. Наибольшее их развитие происходит в районах с влажным климатом.
Церкоспороз столовой и сахарной свеклы, вызываемый церкоспорой свекольной (Cercospora beticola),- одно из наиболее опасных и распространенных заболеваний этих культур. Он известен почти во всех районах свеклосеяния и при массовом развитии вызывает большие потери урожая. Так, по данным Американской ассоциации свекловодов, в некоторых штатах США потери урожая составляют до 7,5 т на 1 га.
На листьях и черешках пораженных растений, а также на стеблях семенников образуются сероватые или беловатые пятна с буровато-коричневой или красноватой каймой, диаметром до 3-6 мм (рис. 236). Во влажную погоду на их нижней стороне развивается сероватый налет, состоящий из неветвящихся, буроватых у основания конидиеносцев с бесцветными, многоклеточными, слегка изогнутыми конидиями (рис. 236). В ткани листа развивается сначала бесцветный мицелий, позднее буреющий и уплотняющийся в строму. Его развитие ограничено только пятном. Клетки каймы пятна обладают повышенной физиологической активностью и служат барьером, препятствующим распространению мицелия гриба в здоровые ткани листа.
Вредоносность заболевания определяется повреждением листовых пластинок и снижением в результате этого их фотосинтетической активности. Кроме того, при отмирании листьев происходит развитие новых листьев, нормальный рост корней задерживается. У больных растений уменьшаются масса корня и содержание в нем сахара, а общие потери в сборе сахара при сильном поражении сахарной свеклы церкоспорозом достигают иногда 50-70%.
Церкоспора свекольная развивается на различных представителях семейства маревых - на свекле, лебеде, мари и др.
Некоторые микологи считают, что конидии гриба зимуют на растительных остатках. Однако их жизнеспособность сохраняется только на сухих листьях, а на влажных в природных условиях они быстро ее теряют. Конидии церкоспоры свекольной служат, вероятно, только для расселения гриба в период вегетации, а зимуют плотные мицелиалыше сплетения в листьях, оставшихся на поверхности почвы. Весной на них образуются, конидии, заражающие растения. Кроме того, источником инфекции могут служить сорные растения из семейства маревых.
Уже через 1-3 недели после заражения растений на пятнах развиваются конидии, заражающие новые растения. При повышенной влажности (более 70%) и теплой погоде в, течение лета паразит успевает образовать несколько поколений конидий.
Церкоспора свекольная поражает преимущественно уже закончившие развитие старые листья, поэтому заболевание проявляется обычно в июне, нарастая до конца вегетации растений. Основной источник инфекщщ в период вегетации - нижние старые листья. При сильном поражении растений в конце июля и в августе происходит отмирание листьев, а иногда и полная гибель ботвы. Сильное развитие болезни наблюдается при чередовании длительных влажных и коротких засушливых периодов. В засушливых районах церкоспороз свеклы развивается слабо.
Церкоспороз винограда вызывают четыре вида из рода церкоспора (С. roesleri, С. sessilis, С. vitis и С. vitiphylla), различающиеся по характеру пятен на листьях, форме и размерам конидий. На пораженных листьях образуются оливково-черноватые пятна с бархатистым оливковым или коричневым налетом конидиального спороношения гриба на нижней стороне. При сильном поражении листья желтеют и опадают. Иногда болезнь распространяется на побеги и ягоды. Биология грибов - возбудителей церкоспороза винограда различна в разных районах виноградарства и зависит от климатических условий. В Средней Азии конидии гриба зимуют на листьях винограда, упавших на поверхность почвы, и на его стеблях, а весной вызывают заражение нижних листьев. С повышением температуры болезнь усиливается и переходит на верхние листья. В некоторых районах эта болезнь по вредоносности стоит на втором месте после оидиума (Uncinula necator). В Армении развитие церкоспороза винограда наблюдается лишь в июле, а максимального развития болезнь достигает только к середине сентября, когда урожай уже созрел, поэтому гриб практически не приносит ущерба.
Практическое значение имеют также церкоспора сельдерея (Cercospora apii) - возбудитель церкоспороза сельдерея и церкоспора кофейная (Cercospora coffeicola), приносящая ущерб плантациям кофе в Латинской Америке, а также другие грибы этого рода.
У грибов рода гельминтоспорий характерна форма конидий (рис. 237, 238). Они темноцветные, прямые или слегка изогнутые, цилиндрические, веретеновидные или обратнобулавовидные, с несколькими поперечными перегородками. Конидии образуются у вершины и по бокам конидиеносцев по типу пороспор.
Рис. 237. Гельминтоспорий (Helminthosporium). Конидиеносец с конидиями
В этот род объединяют очень разнородную группу грибов, отличающихся по морфологии и образу жизни. Поэтому из комплексного, формального рода гельминтоспорий выделяют несколько самостоятельных родов: дрекслера, биполярис и курвулярия.
У одного из видов - гельминтоспория турецкого (Helminthosporium turcicum) - найдена сумчатая стадия типа трихометасферия (Trichometasphaeria).
Грибы рода гельминтоспорий, у которых не обнаружена сумчатая стадия и конидии образуются одновременно на вершине и по бокам конидиеносца, сохранили за собой и название рода. Это преимущественно сапрофиты на коре, листьях, сухих ветвях и стеблях древесных и травянистых растений. Многие из них еще мало изучены и не имеют большого практического значения.
Болезни злаков, вызываемые гельминтоспориями, носят общее название гельминтоспориозов, хотя по форме и проявлению они весьма разнообразны. Гельминтоспориоз может выражаться в виде корневой гнили, различного типа пятнистостей листьев, черных узлов на стеблях, сажистых налетах на колосьях в виде черного зародыша семян. Один и тот же вид паразита может вызывать сразу несколько типов заболеваний. Например, гелъминтоспорий ячменный (Н. sativum) - возбудитель корневой гнили, пятнистости листьев, сажистого налета на колосьях и "черного зародыша" семян пшеницы, ячменя и других злаков. Часто разные виды паразитов вызывают одинаковый типа заболевания. Например, полосатую пятнистость листьев вызывают гельминтоспорий злаковый (Н. gramineum) и гелъминтоспорий овсовый (Н. avenae).
Чтобы представить себе серьезный ущерб, который приносят человечеству гельминтоспориозы злаков, достаточно вспомнить страшный голод в 1943 г. в Бенгалии (Индия). Он был вызван гибелью посевов риса в 1942 г. от гельмиотоспория рисового (Н. oryzae). Один из индидийских микологов, направленный на работу в Бенгалию в это время, так описывает свои впечатления. На всем пути через Бенгалию до города Дакка лежали мертвые или голодные и умирающие люди. Голод продолжался несколько месяцев и унес 2 млн. жизней. Эпифитотия гельминтоспориоза в Бенгалии по своей силе и количеству человеческих жертв может сравниться только с картофельным голодом в Ирландии в 1845 г., вызванным эпифитотией фитофтороза.
Такие сильные эпифитотии, приводящие к гибели людей от голода, к счастью, бывают редко. Однако ежегодные потери от гельминтоспориоза зерновых культур, являющихся основными продовольственными культурами мира, все же огромны.
При заболевании злаков корневой гнилью поражаются первичные и вторичные корни, подземное междоузлие и основание стебля. На пораженном органе появляются сначала светло-бурые пятна или полоски, которые постепенно темнеют, сливаются и захватывают значительную часть органа или весь орган. В результате загнивания корней и основания стебля происходит отставание растения в росте, отмирание продуктивных стеблей в фазе цветения или молочной спелости, недоразвитие зерна в колосьях, а иногда и полная потеря зерна (белоколосость).
Для возбудителей корневой гнили характерна широкая специализация, способность поражать большой круг растений-хозяев. Главным возбудителем корневой гнили является гелъминтоспорий ячменный (Н. sativum, табл. 61.).

Таблица 61. Гифомицеты и меланкониевые грибы - паразиты растений: 1 - кладоспориоз томатов (возбудитель - Cladosporium fulvum), внизу - спороношение гриба; 2 - альтернариоз капусты (возбудитель - Alternaria brassicae), внизу - конидии паразита; 3 - антракноз фасоли (возбудитель - Colletorichum lindemuthianum), рядом-спороношение гриба; 4 - антракноз тыквы (возбудитель - С. lagenarium), справа - спороношение гриба; 5 - антракноз винограда (возбудитель - Gloeosporium ampelinum); 6 - гельминтоспориоз ячменя (возбудитель - Helminthosporium sativum), слева - спороношение гриба
Гельминтоспорий вызывает заболевание зерна, которое проявляется в виде скрытой инфекции или в виде "черного зародыша" (потемнения зерна). В последнем случае черный налет гриба хорошо заметен на зерне (рис. 238). Больные семена теряют всхожесть. Поражение семян вызывают почти все гельминтоспории, так как сохранение инфекции с семенами - надежный способ сохранения и зимовки возбудителя в природе.
Рис. 238. Гельминтоспорий (Helminthosporium): 1 - пораженная зерновка ячменя; 2 - конидии
Гельминтоспорий вызывает несколько типов поражения листьев, чаще всего это полосатая, глазковая и сетчатая пятнистости. В пораженных листьях отмирает ткань и листовая пластинка буреет, засыхает и расщепляется. Такой тип поражения характерен для гелъминтоспория злакового (Н. gramineum). У гелъминтоспория викториц (Н. victoriae) полосчатость сопровождается покраснением и обесцвечиванием листа.
При сетчатой пятнистости развиваются продолговатые небольшие бурые пятна, разбросанные по листовой пластинке.
Глазковая пятнистость - это светлые овальные пятна, окаймленные темно-коричневым ободком. Ее вызывают гелъминтоспорий ячменный (Н. sativum) и другие виды. При поражении листьев почти всегда на пятнах развивается темный налет спороношения патогена. Пятна обычно переходят в некроз.
У некоторых гельминтоспориев в развитии листовых симптомов болезни принимают участие токсины, выделяемые патогеном. Образование токсинов пока обнаружено только у шести видов рода. Образуемые гельминтоспориями токсические соединения оказывают влияние или нарушают жизненно важные функции растения, такие, как проницаемость клеточных мембран, механизмы гормонального контроля роста, фиксация СО2, дыхание, окислительное фосфорилирование, фенольный синтез и другие процессы. Нарушение этих функций ослабляет растение, облегчает грибу захват новых тканей и приводит к появлению характерных симптомов болезни, механизмы этих процессов пока еще детально не изучены. Предполагают, что специфические токсины, как викторин (Н. victoriae), кохлиоболин (Н. oryzae), карботоксин (Н. carbonum), являются главным фактором, определяющим развитие заболевания. Эти токсины у гельминтоспориев имеют пептидную природу.
Неспецифические токсины обычно действуют позднее в процессе развития болезни, и, по-видимому, их действие приводит к появлению симптомов болезни. Два неспецифических токсина терпеноидной природы (гельминтоспорал и гельминтоспорол) обнаружены у гельминтоспория ячменного.
Гельминтоспориям свойственна большая изменчивость и быстрое приспособление к новым сортам растений-хозяев. Например, недавно появилась раса "Т" у гельминтоспория кукурузного (Н. maydis). Этот гриб вызывает широко распространенное заболевание кукурузы - южный гельминтоспориоз. В 1970 г. эта болезнь кукурузы охватила несколько штатов США. В том же году болезнь была обнаружена в Югославии на кукурузе, завезенной из США. Поля кукурузы выглядели обожженными. При высокой температуре и достаточной влажности весь посев погибал за 10-14 дней. Кукурузоводство встало перед серьезной проблемой. Оказалось, новая раса гриба поражала только гибриды с техасской цитоплазменной мужской стерильностью, которые широко введены в культуру в США. "Т"-ген мужской стерильности избавил людей от дорогостоящего ручного труда по удалению мужских соцветий для получения гибридов при скрещивании, но дал растения, восприимчивые к новой расе гриба. Формирование этой расы шло в 1963-1969 гг., т. е. с момента появления в культуре новых гибридов.
Наиболее распространены следующие виды гельминтоспория.
Гельминтоспорий ячменный имеет сумчатую стадию кохлиоболюс ячменный (Cochliobolus sativus). Он поражает ячмень, пшеницу, рожь и 83 вида дикорастущих трав. Наибольшей восприимчивостью из них обладают пырей, костер, мятлик, овсяница и некоторые другие. Все эти злаки являются резервуарами патогена в природе. Гриб способен поражать все органы растения-хозяина. Он вызывает корневую гниль, пятнистость листьев, "черный зародыш", сажистые налеты на колосьях, поражает всходы. Сохраняется на растительных остатках и передается с семенами.
Гельминтоспорий злаковый имеет сумчатую стадию пиренофора злаковая (Pyrenophora graminea). Этот вид - космополитный. Он вызывает полосатую пятнистость листьев ячменя. Поражает семена и всходы. В цикле его развития наблюдается несколько стадий: конидиеносцы с конидиями - летом, склероции на остатках больных растений - зимой, аскостромы на растительных остатках - весной. Патоген сохраняется в виде склероциев и мицелия (в семенах).
Гельминтоспорий овсовый имеет сумчатую стадию - пиренофора овсовая (Pyrenophora avenae). Она вызывает гибель всходов и полосатую пятнистость листьев овса. Инфекция сохраняется на семенах и растительных остатках.
Гельминтоспорий рисовый имеет сумчатую стадию - кохлиоболюс миябеанус (Cochliobolus mijabeanus). Гриб вызывает выпадение всходов, глазковую пятнистость листьев и черную точечность колосковых чешуй риса. Инфекция сохраняется с семенами и на растительных остатках. Главная область распространения - страны Юго-Восточной Азии.
Гельминтоспорий турецкий имеет сумчатую стадию, трихометасферия турецкая (Trichometasphaeria turcica). Гриб поражает листья, початки и (редко) корневую шейку кукурузы. Вредоносность гриба заключается в том, что он способен вызывать преждевременное засыхание листьев кукурузы в период формирования початков. Инфекция сохраняется в семенах и на растительных остатках. Заболевание описано во всех странах, где возделывают кукурузу.
У грибов этого рода многоклеточные темно-окрашенные конидии с поперечными и продольными перегородками. Форма конидий разнообразна и представляет собой вариации формы яйцевидного типа. Верхний конец конидии вытянут в короткий или длинный "носик". У многих альтернарий конидии образуют легко распадающиеся цепочки (рис. 239). Однако среди альтернарий есть представители с одиночно сидящими конидиями, у которых "носик" обычно вытянут в длинную нить. Конидиеносцы всегда темно-окрашенные, простые или у вершины ступенчато-изогнутые.
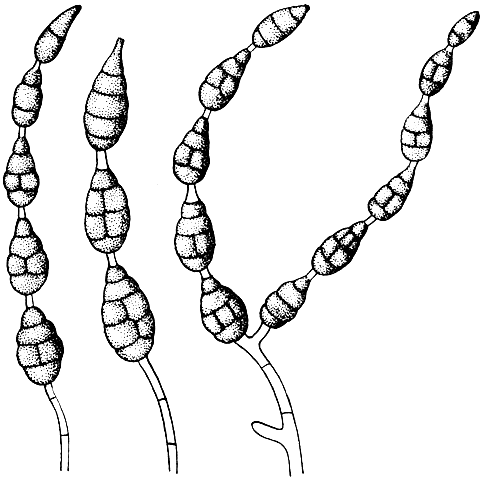
Рис. 239. Альтернария (Alternaria). Конидиеносец с конидиями
Замедленная киносъемка процесса образования цепочки конидий у альтернарии дала возможность отметить несколько интересных моментов, происходящих при формировании конидии. Обычно молодая конидия появляется на вершине конидиеносца или предыдущей конидии в виде сферического образования. Через 10 мин после появления она начинает вытягиваться и увеличивать свой объем. Приблизительно через 2,5 ч конидия достигает окончательного объема. Новая конидия появляется на ее вершине только после образования первой поперечной перегородки. Каждый раз перед образованием новой конидии наблюдается резкое уменьшение объема уже имеющихся в цепочке спор. Уменьшение объема идет от нижней споры к верхней. После образования конидии объем медленно нарастает и опять резко падает перед формированием следующей конидии в цепочке. Такие колебания объема указывают на то, что в цепочке конидий происходит как бы "отсасывание" цитоплазмы к вершине цепочки в момент формирования молодой конидии.
В связи с тем что многие виды альтернарий вызывают заболевания важных сельскохозяйственных культур, потребовалось тщательное изучение окружающих условий, влияющих на спороношение гриба. Оказалось, у альтерпарии пасленовой (A. solani) для образования на мицелии конидиеносцев и формирования конидий нужны разные условия. Влажность и свет являются главными факторами, способствующими появлению конидиеносцев. Для того чтобы на конидиеносцах стали образовываться конидии, нужны пониженная температура и темнота. Следовательно, влияние погодных условий может ускорить или замедлить переход гриба из одной фазы развития в другую и ускорить или замедлить жизненный цикл патогена, т. е. повлиять на развитие болезни, вызванной альтернарией. Человек, зная все фазы развития патогенного гриба и условия, способствующие прохождению этих фаз, может, воздействуя в определенный период на гриб, повлиять на ход развития болезни. Знание всех фаз развития гриба позволяет также предсказывать степень развития болезни в разных климатических условиях и бороться с ней. Развитие эпифитотии зависит от продолжительности сменяющих друг друга периодов.
Альтернарии широко представлены в природе. Многие из них - сапрофиты и развиваются на любых органических субстратах. Резервуаром альтернарий являются отмирающие растения и растительные остатки, с которых гриб попадает в почву. Наряду с другими грибами альтернария принимает участие в разложении и минерализации растительных остатков. Этому способствует огромный комплекс ферментов, обнаруженный у сапрофитных альтернарий.
Сапрофитные виды альтернарий, обладающие высокоактивной полигалактуроназой, вызывают размягчение огурцов при посоле, разлагают глюкозид рутин, который содержится в кожуре плодов яблок, листьях чая, табака и других растений, придавая им желто-оранжевую окраску.
Богатый ферментный аппарат гриба обеспечивает широкую амплитуду приспособленности и способность существовать в достаточно разнообразных условиях. Этому также благоприятствует легкое распространение спор ветром. Споры альтернарии, иногда даже соединенные в цепочки, обнаруживают в воздушных массах везде, где есть растения.
Некоторые сапрофитные виды, например альтернария тонкая (A. tenuis), альтернария тончайшая (A. tenuissima), альтернария кочанно-капустная (A. oleracea), при сильном ослаблении растений могут проявлять паразитические свойства. Они поселяются на старых листьях, а с них переходят на стебель и молодые листья. Это наблюдают особенно часто в случаях поражения растения каким-нибудь другим грибом: альтернария проявляется тогда уже в качестве вторичного паразита.
Интересно отметить, что эти же сапрофитные виды альтернарий обычно встречаются на листьях здоровых растений в качестве эпифитов. Там они развиты слабо, так как располагают ограниченным количеством питательных веществ, присутствующих на листовой поверхности, и подавляются специфическими веществами, выделяемыми растением. Можно полагать, что при ослаблении растения они получают шанс для более интенсивного развития. Пока это свойство эпифитных микроорганизмов недостаточно изучено, но полезные свойства эпифитной микофлоры уже отмечены. Эпифитные организмы способны подавлять развитие патогенов, попадающих на листья. Это установлено на примере альтернарий.
Эпифит альтернария тончайшая (A. tenuissima) подавляет развитие возбудителя болезни цинний - альтернарии цинниевой (A. zinniae). Здесь происходит не только прямое подавление прорастания спор патогена под влиянием выделений эпифита, но, одновременно с этим, эпифит стимулирует выделение ингибирующих веществ самого растения.
Альтернарии всегда присутствуют на семенах растений. При сильном развитии гриба семя теряет всхожесть. Во многих случаях присутствие гриба не сказывается на дальнейшем развитии растения. Однако грибы некоторых видов альтернарий сохраняют и передают инфекцию с семенами, например алыперпарии капустная (A. brassicae, табл. 61), морковная (A. dauci), гвоздичная (A. dianthicola), цинниевая (A. zinniae) и др. В этих случаях гриб поражает проростки либо инфекция сохраняется на корневой шейке и обнаруживает себя в момент цветения или плодоношения, когда устойчивость растения снижается. Если условия для развития растений неблагоприятны, то инфекция проявляется раньше стадии зрелости растения. Возбудитель пятнистости хлопчатника (рис. 240), цитрусовых и многих других культур - алътернария тонкая (A. tenuis) - образует тентоксин. При вымачивании семян хлопчатника и других растений в тентоксине наблюдается сильный хлороз проростков как результат ингибирования токсином процесса образования хлорофилла.

Рис. 240. Альтернариоз хлопчатника
В связи с поисками биологических методов борьбы с болезнями растений представляет интерес алътернария повиликовая (A. cuscutocidae). Она вызывает побурение и гибель стеблей повилики, распространенного паразита многих культурных растений. Из-за тесного контакта повилики с растением-хозяином трудно применять химические методы борьбы. Опрыскивание люцерновой повилики спорами альтернарии в Киргизии дало положительные результаты.
В настоящее время известно более 100 видов альтернарий, из них только около 25 наиболее широко распространены и имеют практическое значение.
Алътернария пасленовая (A. solani, табл. 61) поражает картофель и томаты. Болезнь возникает в период бутонизации и приводит к преждевременной гибели ботвы. На листьях появляются концентрические сухие пятна со спороношением гриба. Гриб поражает плоды и клубни, на которых образуются сухие темные пятна. Споры, крупные, с длинным "носиком", сидят на конидиеносцах одиночно. Гриб зимует в пораженной ботве, встречается в почве.
Альтернария капустная наносит наибольший вред семенной капусте. На стручках и листьях образуются пурпурные, позднее темнеющие пятна, на которых появляется спороношение гриба. Конидии одиночные или в коротких цепочках. Семена капусты гибнут или дают низкую всхожесть. Семенная инфекция служит источником заражения почвы и причиной гибели проростков.
Этот род близок к роду альтернария как по образу жизни его представителей, так и морфологически (у его представителей тоже многоклеточные темноокрашенные конидии). Однако конидии здесь никогда не образуют цепочек и не имеют "носика" - вытянутой верхушечной клетки. Самыми характерными чертами этого рода являются строение и способ пролиферации конидиеносцев. Клетка конидиеносца, несущая конидию, вздута у вершины. После образования первой конидии через вершину вздутия конидиеносец продолжает расти, отодвигая первую конидию в сторону. После некоторого роста вершина конидиеносца снова вздувается, образуется следующая конидия, отодвигаемая в сторону новым выростом конидиеносца, и т. д. Обычно происходит три и более пролифераций, и на конидиеносце кистевидно располагаются 3-5 конидий (рис. 241).
Рис. 241. Стемфилий (Stemphylium). Конидиеносец с конидиями
Молодые конидии стемфилия округлые, яйцевидные или обратнояйцевидные (зрелые сохраняют эту форму или становятся широкоэллиптическими либо несколько удлиненными). Оболочка конидий гладкая или шиповатая. Место прикрепления конидии бывает окружено зоной усиленной пигментации. Размеры конидий достигают нескольких десятков микрон.
Сумчатая стадия известна у шести видов рода и относится к роду плеоспора (Pleospora).
Стемфилий - космополит и встречается на растительных остатках, семенах различных растений, бумаге, на зрелых плодах при транспортировке и хранении. В подавляющем большинстве это сапрофиты, но некоторые виды можно назвать факультативными паразитами. Они вызывают пятнистость на листьях и стеблях растений, являются причиной гибели семян.
Цикл развития таких слабых паразитов в общих чертах следующий. В течение долгого времени гриб может быть сапрофитом на отмерших растительных остатках, откуда он попадает в почву и на листья растений. При попадании спор или мицелия гриба на ослабленные растения происходит заражение самых нижних и старых листьев. На листьях появляются пятна, которые постепенно распространяются на более молодые части растения.
Особенно легко заражаются цветущие и плодоносящие растения, устойчивость которых значительно снижена.
Распространение паразита по растению заканчивается заражением плодов и семян. На семенах гриб может оставаться живым в течение нескольких лет. В зависимости от степени поражения семени оно теряет всхожесть или дает проросток, на котором сохраняется инфекция. Сохраняющаяся на корневой шейке инфекция проявляет себя на старых Листьях в момент цветения или плодоношения растения. Инфекция может проявить себя раньше, если растение ослаблено какими-нибудь неблагоприятными факторами. Такими факторами могут быть погодные условия, повреждение растения или заражение его другими патогенными организмами. Например, стемфилий часто встречается на луке, зараженном пероноспорой. Но это не значит, что стемфилий не может заразить лук без пероноспоры. Если растения лука были ослаблены неблагоприятными погодными условиями, то на них происходит массовое развитие гриба. Наиболее распространены 2 вида стемфилия.
Стемфилий обилънокистевой (S. botryosum) встречается на растительных остатках и семенах многих растений, хорошо растет на бумаге и соломе льна, делая солому ломкой. Этот гриб образует гниль на зрелых плодах томатов и яблоках. Обнаружен на 64 видах растений в качестве слабого паразита. Чаще других растений поражает салат, лук и люцерну (особенно в условиях повышенной влажности). Гифы гриба проникают внутрь листьев через устьица растения, и уже через 120 ч на листьях появляются споры гриба. Конидиеносцы имеют от 1 до 4 последовательных вздутий. Конидии округло-квадратные, яйцевидные, обычно шиповатые, суженные у центральной поперечной перегородки. Сумчатая стадия гриба - плеоспора травяная (P. herbarum) - встречается на растительных остатках после перезимовки и в культуре гриба в лаборатории при пониженной температуре. Сумки имеют двойную оболочку и содержат по восемь аскоспор такого же типа, как и конидии гриба, но цилиндрической или булавовидной формы.
Стемфилий capцинообразный (S. sarciniforme), в отличие от предыдущего вида, встречается на более ограниченном круге растений-хозяев, принадлежащих преимущественно к семейству бобовых. Гриб вызывает пятнистость листьев красного и белого клевера, внедряется в клетки растения через эпидермальные клетки.
Сумчатая стадия стемфилия сарцинообразного не найдена. Конидии более округлой формы, чем у стемфилия обильнокистевого, с гладкой оболочкой и без сужения у поперечной перегородки.
У многих видов этого рода имеются только конидиальные спороношения и отсутствует половая стадия. (У других известны половые совершенные стадии.)
Конидиальное спороношение у фузариумов чрезвычайно разнообразно по морфологии (табл. 60) и способу образования конидий. Грибы этого рода имеют два типа конидий - макро- и микроконидии.

Таблица 60. Поражение растений фузариумом: 1 - фузариозное увядание льна (возбудитель - Fusarium oxysporum f. lini); 2 - фузариоз колосьев пшеницы (возбудитель - F. graminearum), вверху - конидии паразита; 3 - сумчатая стадия гриба (Gibberella zeae) на колосках пшеницы; 4 - сумки этого гриба со спорами; 5 - перитеции этого гриба; 6 - снежная плесень озимой ржи (возбудитель - Fusarium nivale); 7 - сумки возбудителя заболевания (Micronectriella nivalis); 8 - перитеции этого гриба
Макроконидии образуются в воздушном мицелии на простых или ветвящихся конидиеносцах (рис. 242), или представляют собой скопления массы конидиеносцев в виде спородохиев, или образуют пионоты. Макроконидии большей частью с 3-5 перегородками, реже с 6-9, веретеновидные, веретеновидно-серповидные, серповидные, реже ланцетовидные. У основания конидий имеется четко или слабовыраженная ножка, или конидии лишены ее. Верхняя клетка макроконидий имеет характерную форму: она удлиненная или заостренная, может быть тупая или клювовидная и другой формы. В массе макроконидии светлоокрашенные (бело-охряные, охряно-розовые, оранжевые, синие, сине-зеленые).
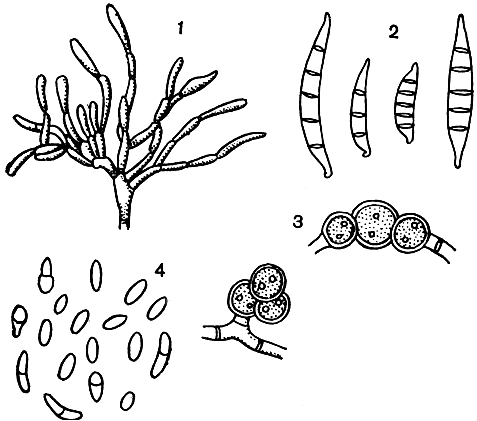
Рис. 242. Спороношение фузариума (Fusarium): 1 - формирование конидий; 2 - макроконидии; 3 - хламидоспоры; 4 - микроконидии
Микроконидии образуются обычно в воздушном мицелии на простых или сложных конидиеносцах, в цепочках или собраны в головки, а также часто в виде скоплений между гифами мицелия. Конидии одноклетные, реже с одной и еще реже с 2-3 перегородками, обычно овальные, яйцевидные, эллипсовидные, реже шаровидные, грушевидные и веретеновидные.
Мицелий у этих грибов чаще белый, бело- розовый, розово-сиреневый или бурый.
Кроме того, у отдельных видов рода фузариум в мицелии (а иногда в конидиях) образуются хламидоспоры - одноклетные части гиф, обособляющиеся от остальных клеток толстой оболочкой. Последние обычно бесцветные, реже окрашены в охряно-коричневые тона. Могут быть единичные или собраны в узлы, иногда в виде цепочек (рис. 242).
В некоторых случаях у этих грибов образуются склероции - тесное скопление гиф роговидной консистенции беловатого, желтоватого, коричневого или синего цвета. Это покоящаяся стадия. Развиваются они чаще всего в тканях субстрата или в почве и служат у этих грибов, как и хламидоспоры, для перезимовки и перенесения неблагоприятных условий, способствуя тем самым сохранению вида.
Грибы рода фузариум широко распространены в природе. Виды этого рода, как и другие микроскопические грибы, участвуют в общем круговороте веществ в природе. Обладая большим разнообразием ферментов, они способны разрушать органические соединения. Отдельные виды рода фузариум обладают свойством синтезировать различные биологически активные вещества (например, витамины, антибиотики и токсины). Грибы этого рода развиваются преимущественно в почве, однако могут существовать на различных растительных и животных субстратах в воде и других местах обитания.
Фузариум сохраняется в почве, на растительных остатках, а частично и в самих растениях в виде мицелия, хламидоспор, а в случае наличия сумчатой стадии - в виде перитециев. Конидии этого гриба могут переноситься водой, насекомыми, орудиями производства и воздушными течениями. Лучшее развитие грибов из рода фузариум, как и многих других, происходит при повышенной влажности воздуха и почвы. Немаловажным фактором для их существования является также тепло. Сочетание необходимой температуры и влажности обусловливает массовое развитие этих грибов.
Род фузариум в целом представляет обширную биологически неоднородную группу грибов. Среди них есть и достаточно выраженные паразиты растений, и группы полупаразитов, способных поражать только ослабленные организмы, и, наконец, сапрофиты, живущие на растительных остатках, в почве и на отмерших частях растений. Известны фузариумы, паразитирующие на насекомых, а также вызывающие микозы и токсикозы человека и тепло-кровных животных.
Большинство грибов этого рода - фитотрофы (возбудители заболеваний растений). Фузариумы одного вида могут поражать растения из самых разнообразных семейств, вызывая у них различные патологические явления - гниль корней, семян, плодов, а также общее угнетение и преждевременное увядание. Тип проявления болезни зависит, с одной стороны, от видовой принадлежности патогенов, а с другой, от экологических условий, в которых они развивались. К наиболее распространенным фузариозным заболеваниям относятся гниль корней и трахеомикозное сосудистое увядание растений.
Разнообразная гниль корней сильнее всего проявляется в фазу всходов, но может также развиваться в течение всей вегетации растений. У пораженных всходов происходит побурение и загнивание корней, а также утончение прикорневой части стебля. Часто болезнь поражает и семядоли, на них образуются вдавленные красно-бурые пятна, которые (при повышенной влажности) покрываются белым или бело-розовым налетом гриба. Подобную картину можно наблюдать при поражении всходов люпина, сои, фасоли и других бобовых культур. Часто гибель всходов происходит еще до выхода их на поверхность. При заражении растений в более поздние фазы наблюдается отставание растений в росте и развитии. У злаков, кроме загнивания корней, часто происходит отмирание стеблей или наблюдается пустоколосость. Кроме того, пораженные корневой гнилью растения, как правило, легко выдергиваются из почвы. Заболевание встречается повсеместно, но наиболее интенсивно развивается на участках с тяжелыми почвами, а также в местах с избыточным увлажнением.
Наиболее распространенными возбудителями корневой гнили многих культур (например, гороха, фасоли, огурцов, дыни, арбузов, томатов) являются такие виды, как F. avenaceum, F. solani, F. culmorum, F. gibbosum, F. semitectum, F. javanicum, F. heterosporum и др. Один из основных возбудителей корневой гнили злаков - гриб F. graminearum. В северных районах возделывания хлебных злаков широко распространено заболевание под названием "снежная плесень", вызываемое грибом F. nivale (табл. 60). Болезнь приводит к выпреванию хлебов. В ряде случаев заболевание корней носит комплексный характер, т. е. наряду с фузариозной имеется бактериальная и вирусная инфекция.
Трахеомикозное увядание растений проявляется в период всходов, но чаще болезнь поражает растения на более поздних фазах их развития. Заболевание встречается преимущественно в районах с сухим и жарким летом. Наиболее характерно для этой болезни у большинства культур - поникание верхушки растений, потеря тургора листьев, их пожелтение, увядание и полное усыхание. Увядание часто проявляется очагами. При поражении растений до цветения они задерживаются в росте и развитии, при этом болезнь может развиваться сравнительно медленно (хроническая форма). На более поздних фазах при повышенной температуре и недостатке влаги она может протекать более интенсивно и гибель растений наступает очень быстро (в течение 6-7 дней).
Гибель растений, пораженных трахеомикозным увяданием, происходит преимущественно за счет токсинов, а также вследствие закупорки проводящих сосудов гифами гриба. На поперечном срезе пораженного стебля наблюдается побурение сосудистой системы. Гриб способен распространяться по сосудам вверх. Больные растения легко выдергиваются из почвы, но основной корень остается, как правило, внешне здоровым, тогда как мелкие корешки буреют и загнивают.
Пораженные трахеомикозным увяданием растения обычно не образуют плодов и зерновок либо имеют недоразвитые плоды и зерновки со щуплыми семенами.
Заболевание чрезвычайно широко распространено и поражает самые разнообразные культуры (пшеница, рожь, рис, бобы, горох, соя, люцерна, дыня, арбуз). Его также считают одним из вредоноснейших заболеваний льна и хлопчатника. У последнего при поражении, кроме симптомов, описанных выше, на листьях образуются желтые жилки, а участки тканей между ними остаются зелеными, отчего лист становится сетчатым. Фузариозное увядание картофеля также довольно распространенное заболевание на юге и юго-востоке нашей страны.
В закрытом грунте от этого заболевания часто погибает рассада капусты и томатов. В теплицах и парниках известны также трахеомикозные заболевания многих цветочных растений (астр, гвоздики, левкоев, цикламенов и др.). В питомниках от фузариозного увядания погибают сеянцы хвойных пород.
Возбудитель трахеомикозного увядания - гриб F. oxysporum и его многочисленные формы.
Болезни плодов и семян, вызываемые фузариумом, причиняют большой вред. У большинства культур заражение происходит в тот период, когда растения находятся еще на корню, и чаще всего в районах с влажной второй половиной лета или в годы с повышенным количеством осадков в тот же период. В основном это бывает у зерновых и бобовых культур. Например, при поражении злаков грибом Fusarium graminearum колоски приобретают белесоватую окраску и на них развиваются розовые или оранжевые подушечки, состоящие из спороношения гриба. Через некоторое время на пораженных колосках появляются темные скопления плодовых тел (перитециев), представляющих собой сумчатую стадию возбудителя - Gibberella zeae. Подобную картину можно наблюдать и при поражении зерновок риса F. moniliforme с сумчатой стадией G. fujikuroi. Пораженное зерно становится щуплым, теряет всхожесть, приобретает ядовитые свойства, его нельзя употреблять в пищу.
Заражение семян бобовых культур также чаще происходит в полевых условиях в момент созревания плодов, которые являются благоприятным субстратом для развития патогена. На створках бобов появляется белый, бело-розовый налет гриба. При повышенной влажности его мицелий проникает внутрь бобов, поражая семена. Последние становятся щуплыми, теряют всхожесть или дают больные проростки (рис. 243, 244). Такие семена служат источником инфекции и приводят к гибели всходы и взрослые растения.
Рис. 243. Поражение семян люпина фузариумом

Рис. 244. Поражение проростка кормовых бобов
Развитие фузариозов у естественно зараженных семян может продолжаться при хранении в условиях повышенной влажности, причем здесь происходит и перезаражение. Поражение семян вызывают все грибы рода фузариум. Особенно часто на бобовых встречаются виды F. avenaceum, F. solani, F. culmorum, F. oxysporum. F. sporotrichiella встречается на семенах бобовых культур довольно редко, но он заслуживает особого внимания, так как этот патоген может оказывать токсическое действие на организм человека и животных.
Некоторые грибы рода фузариум поражают коробочки плодов. Подобное явление наблюдают на льне, хлопчатнике, кенафе и конопле. Семена льна становятся ядовитыми, и из них получается так называемое "пьяное масло", а при поражении коробочек хлопчатника и других технических культур происходит загнивание как семян, так и содержащегося в коробочках волокна. Это приводит к снижению качества и количества урожая.
Часто наблюдается поражение семян, плодов и корнеплодов грибами этого рода в условиях хранения. К широко распространенным заболеваниям, развивающимся во время хранения, относятся кагатная гниль свеклы и сухая гниль картофеля. Кагатная гниль проявляется в отмирании и разложении тканей корня, которые теряют сахар и становятся совершенно непригодными для производства. Возбудитель заболевания - гриб F. culmorum. Сухая гниль картофеля характеризуется образованием на клубнях бурых пятен, слегка вдавленных позднее, на поверхности которых образуются выпуклые подушечки белого или кремового цвета, состоящие из грибницы и большого количества спор гриба. Чаще всего возбудителями бывают F. solani, F. solani var. coeruleum и F. avenaceum.
Интоксикация сельскохозяйственных продуктов, вызванная фузариумом, может стать причиной отравления людей и животных. Заболевание людей, связанное с употреблением в пищу зерна с токсическими свойствами, называют "пьяным хлебом". Отравление людей сопровождается головными болями, головокружением, общей слабостью и расстройством желудка. Причина заболевания - гриб F. graminearum и некоторые близкие к нему виды (например, F. avenaceum и F. nivale). Они развиваются на хлебных злаках еще в поле. Заражение озимых происходит от инфекции, находящейся в почве в мицелии или на остатках растений в виде мицелия, конидий или сумкоспор в перитециях. Токсические свойства приобретают семена всех хлебных и кормовых злаков. Симптомы отравления часто наблюдаются и у многих домашних животных при поедании ими токсичного зерна или корма.
Заболевание людей алиментарно-токсической алейкией также связано с употреблением в пищу перезимовавшего в поле зерна, пораженного грибом F. sporotrichiella var. sporotrichioides. После перезимовки токсичными свойствами обладают все хлебные злаки, но особенно просо. У людей, работающих с таким зерном, наблюдается раздражение слизистой оболочки носовой полости и верхних дыхательных путей.
Часто это заболевание сопровождается кровотечением из носа, чиханием и кашлем. Скармливание такого зерна лошадям, крупному рогатому скоту приводит к их отравлению и даже к гибели.
Известны также фузариумы, паразитирующие на грибах. Например, F. sporotrichiella успешно развивается на ржавчинных и головневых грибах.
Фузариум может быть возбудителем кожных заболеваний на человеке, вызывая всевозможные виды дерматитов.
Известны представители рода фузариум как грибы, поселяющиеся на книгах и вызывающие их порчу. Некоторые из фузариумов развиваются и на кожаных изделиях. Кроме того, среди многочисленных микроорганизмов, вызывающих загрязнение воды, встречается также фузариум.
Микоризные грибы играют положительную роль в питании высших растений. Наряду с широко распространенным представлением о фузариумах как о факультативных паразитах, высказывается мнение об их фитосимбиотрофной природе и принадлежности к микоризным грибам. Некоторые виды рода фузариум развиваются в корневой зоне растений или на их корнях и не всегда обладают паразитическими свойствами. Эти грибы накапливаются в почве при длительном выращивании растений и используют в качестве питания не только органические вещества почвы, но и корневые выделения растений. Находясь в тесном контакте с клетками корня, фузариум усваивает их питательные вещества и в то же время предоставляет растению соединения, необходимые для его развития. Таким образом, тот фузариум, который развивается в тканях корней определенных растений и не вызывает губительного действия на рост и развитие последних, может быть отнесен к микоризным или фитосимбионтным грибам. Например, грибы F. sambucinum и F. heterosporum, развивающиеся на корнях пшеницы, оказывают положительное влияние на рост и развитие последней и особенно на ее корневую систему. Многие представители рода фузариум являются лишь спутниками возбудителей болезней. Так из многочисленных видов грибов этого рода, участвующих в поражении свеклы кагатной гнилью, не все являются патогенными.
Грибы рода фузариум могут продуцировать биологически активные вещества, которые применяют в сельском хозяйстве. Например, свойство гриба F. moniliforme оказывать стимулирующее действие на рост растений было использовано для получения стимулятора роста - гиббереллина.
Грибы рода Fusarium часто образуют антибиотики.
Водные гифомицеты (И. А. Дудка)
Водные гифомицеты являются составной частью экологической группы водных грибов, куда, помимо представителей несовершенных грибов, входят также представители классов хитридиомицетов (Chytridiomycetes), оомицетов (Oomycetes), сумчатых (Ascomycetes) и базидиальных (Basidiomycetes) грибов.
Отдельные виды водных гифомицетов были известны еще с конца XIX в. (Tetracladium marchalianum, Anguillospora longissima, Lemonniera aquatica, Clavariopsis aquatica и др.), хотя их во многих случаях принимали за водоросли. Как группа, насчитывающая несколько десятков видов и повсеместно распространенная в водоемах различных типов, водные гифомицеты были открыты английским микологом Ц. Инголдом только в 1940 г. Исследуя под микроскопом каплю воды из пробы, отобранной в июле в 1940 г. в небольшом ручье возле города Лейчестера (Англия), Инголд обнаружил в ней споры двадцати различных видов, среди которых преобладали необычные, бесцветные, четырехлучевые. Но происхождение этих спор оставалось невыясненным: попали ли они в водоем из воздуха, или образовались на каком-либо из субстратов, погруженных в воду. И лишь в ноябре 1940 г. в результате тщательного просмотра всех извлеченных из водоема субстратов было обнаружено, что на гниющих в воде листьях ольхи обильно развиваются типичные для гифомицетов конидиеносцы, на которых образуются многочисленные конидии, по форме идентичные спорам, обнаруженным ранее в воде ручья. Начиная с 1942 г., ученые интенсивно изучают водные гифомицеты в водоемах всех (кроме Антарктиды) континентов земного шара.
Исследования показывают, что водные гифомицеты развиваются преимущественно на погруженных в воду гниющих, так называемых "скелетонизированных" листьях не только ольхи, но и практически всех древесных и кустарниковых листопадных пород. Долгое время считали, что эти грибы не развиваются на погруженных в воду остатках хвойных деревьев. Теперь стали известны случаи нахождения около 20 видов водных гифомицетов на веточках псевдотсуги (Pseudotsuga sp.) и на хвоинках сосны обыкновенной (Pinus silvestris), гниющих в воде ручьев. Вообще погруженные в воду субстраты, на которых развиваются водные гифомицеты, весьма разнообразны: это остатки высших водных растений и веточки различных древесных пород, газетная бумага и косточки слив, чешуйки лука, клубни картофеля и т. д. Наконец, совсем недавно водный гифомицет из рода тетракладий Tetracladium marchalianum был обнаружен на икре пеляди (Coregonus peled) в аппаратах Вейса для инкубации икры.
Водные гифомицеты распространены в водоемах различных типов: реках, ручьях, озерах, прудах, причем встречаются они не только в пресноводных, но и в солоноводных водоемах (соленых озерах, эстуариях), но значительно реже.
Наиболее благоприятным местообитанием для водных гифомицетов служат хорошо аэрируемые ручьи лесных долин с довольно быстрым течением воды и с большим количеством скелетонизированных листьев в воде.
Бесцветный септированный мицелий водных гифомицетов развивается в гниющих тканях листа, образуя многочисленные конидиеносцы, которые выходят из листа в воду обычно под прямым углом к его поверхности. Особенно обильно конидиеносцы развиваются на черешке и главных жилках листа. Конидиеносцы водных гифомицетов обычно полностью находятся под водой. Здесь же, под водой, на них образуются необычные по форме бесцветные конидии. Они имеют специфическое четырехлучевое (тетрарадиальное), реже трехлучевое (трирадиальное), многократно разветвленное или сигмоидное строение (рис. 245).
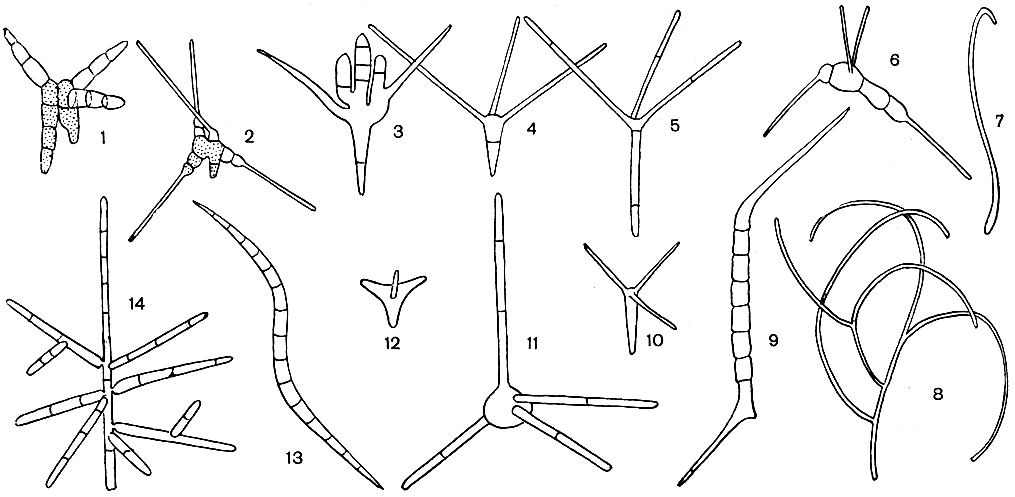
Рис. 245. Конидии водных гифомицетов: 1-6 и 10-12 -четырехлучевые и трехлучевые; 7, 9, 13 - сигмоидные; 8 и 14 - многократно разветвленные
Значительно реже встречаются конидии, имеющие обычное строение, т. е. конидии сферической, эллипсовидной или цилиндрической формы.
По типу онтогенеза четырехлучевые конидии водных гифомицетов принадлежат к алевроспорам. Очень часто у водных гифомицетов четырехлучевые конидии со сходным типом строения отличаются по типу онтогенеза (индивидуального развития) и, наоборот, при одинаковом типе онтогенеза конидии имеют различный четырехлучевой тип строения. Так, у представителей родов клавариопсис (Clavariopsis aquatica) и клаватоспора (Clavatospora longibrachiata) четырехлучевые конидии имеют исключительное сходство в морфологическом строении. У обоих видов они состоят из клиновидной, расширенной на верхушке главной оси, от которой отходят удлиненно-цилиндрические, очень узкие, равномерной ширины по всей длине боковые ответвления. В то же время по типу онтогенеза конидии родов клавариопсис и клаватоспора четко отличаются друг от друга: у первого они являются алевроспорами, у второго - фиалоспорами.
Четырехлучевые споры водных гифомицетов имеют определенное биологическое значение. Есть предположение о двоякой роли четырех-лучевых конидий для выживания водных гифомицетов в условиях водной среды. Споры такого строения дольше парят в сдоях воды, медленнее оседают на дно, в результате чего распространяются на большее расстояние. Такие споры легче закрепляются на подходящих для их развития субстратах, чем споры шаровидной, эллипсовидной или цилиндрической формы, которые легко смываются с субстрата потоком воды. Доказательством значения четырехлучевой, многократно разветвленной и сигмоидной формы конидий для распространения водных гифомицетов в водоемах является тот факт, что конидии именно такой формы преимущественно аккумулируются в своеобразных ловушках - поверхностной пленке и пене рек и ручьев.
Часто можно наблюдать в проточных водоемах, где образуются естественные плотины из камней, упавших в воду стволов деревьев, веток, листьев, обильные скопления пены и пленки. Характерной особенностью этих образований является повышенное поверхностное натяжение, с помощью которого конидии водных гифомицетов улавливаются и задерживаются в пене и пленке. Количество конидий, среди которых преобладают четырехлучевые, многократно разветвленные и сигмоидные, в старой пене, особенно осенью во время максимального развития водных гифомицетов на гниющих в воде листьях, достигает многих десятков тысяч в 1 мл. Конидии, задержавшиеся в пене или пленке от оседания на дно в месте их образования, затем током воды распространяются на некоторое расстояние, в большей или меньшей мере удаленное от места их образования.
Косвенным подтверждением правильности этих гипотез служит обилие спор со щетинками, ответвлениями, разветвленными слизистыми обертками и т. п. у грибов из разных систематических групп, приспособившихся к существованию в пресноводных и соленоводных водоемах.
Аналогичное биологическое значение имеют, по-видимому, и сигмоидные споры водных гифомицетов. Споры такой формы также встречаются у пресноводных и морских сумчатых грибов.
Специфическое строение конидий водных гифомицетов - один из поводов для выяснения возможных путей их возникновения как самостоятельной экологической группы. Предполагают, что водные гифомицеты перешли к существованию в водной среде после продолжительного в историческом понимании существования в наземных условиях, т. е. они являются ремигрантной группой. Такой переход, по мнению Ц. Инголда и В. Кокс, происходил постепенно, главным образом во время осеннего листопада, когда эпифильные наземные гифомицеты вместе с листьями, на которых они развивались, попадали в воду рек, ручьев и других водоемов. Многократное повторение этого процесса привело к возникновению у грибных организмов новых морфологических и физиолого-биохимических при знаков, которые соответствовали требованиям водной среды и способствовали выживанию грибов в этой среде.
К новым морфологическим приспособлениям и особенностям, появившимся у водных гифомицетов, следует отнести длинные ответвления конидий, придающие им вид действительно четырехлучевых и содействующие их выживанию и распространению в водоемах с быстрым течением воды.
В качестве конкретных примеров возможного перехода эпифильных гифомицетов от наземного существования к водному можно рассмотреть два вида: из рода трипосперм (Tripospermum), Т. myrtii, который в наземных условиях развивается как сапрофит на листьях смоковницы (Ficus carica), а во время осеннего листопада конидии его в обилии обнаруживаются в воде ручьев; гриб из рода кампилоспора (Campylospora) С. chaetocladia - типичный водный гифомицет, обнаруженный только в водной среде.
Четырехлучевые конидии кампилоспоры отличаются от конидий трипоспермы только наличием длинных ответвлений, а в остальном строение конидий этих грибов из двух родов очень сходно. Такое сходство конидий послужило основой для предположения о происхождении грибов рода кампилоспора от грибов рода трипосперм в результате постепенного развития у них удлиненных и заостренных на концах ответвлений при попадании конидий трипосперма в воду. По-видимому, аналогичным образом в процессе постепенного перехода от наземного образа жизни к водному возникли и другие водные гифомицеты с их четырехлучевыми конидиями.
Вполне возможно, что окончательное формирование экологической группы водных гифомицетов еще не завершено. Между гифомицетами, весь жизненный цикл которых проходит в водоеме, и гифомицетами, которые развиваются только в наземных условиях существования, имеется множество переходных форм, ведущих амфибийный образ жизни или способных развиваться как в воде, так и на суше. Так, в почве и подстилке были обнаружены представители водных гифомицетов из родов варикоспорий (Varicosporium elodeae), тетракладий (Tetracladium setigerum), алатоспора (Alatospora acuminata), трицеллула (Tricellula aquatica). С другой стороны, на погруженных в воду ручьев гниющих листьях были обнаружены типично наземные, в основном почвенные, гифомицеты из родов фиалоцефала (Phialocephala bactrospora), стахиботрис (Stachybotrys atra), альтернария (Alternaria tenuis) и др. Довольно часто в воде ручьев встречается гифомицет из рода центроспора (Centrospora acerina), сапрофитно развивающийся на погруженных в воду гниющих листьях различных древесных пород. Этот же гриб в наземных условиях развивается на корнеплодах моркови, петрушки и пастернака при их хранении.
Все эти примеры показывают отсутствие четкого разграничения между группами водных и наземных гифомицетов и возможность постепенного перехода некоторых наземных гифомицетов к водному образу жизни и в настоящее время, с образованием специфических для водных гифомицетов четырехлучевых, многократно разветвленных или сигмоидных конидий. Четырехлучевая форма и многократно разветвленная и сигмоидная форма конидий водных гифомицетов оказались наиболее пригодными для осуществления распространения видов в условиях водной толщи. Условия водной среды настолько специфичны и однозначны, что благодаря их влиянию сходные по форме структуры, функцией которых является распространение видов в водоемах, возникли независимо в ходе эволюции разных, часто довольно удаленных друг от друга систематических групп грибов, приспособившихся к существованию в воде.
Для многих несовершенных грибов, в том числе и для гифомицетов, характерно явление плеоморфизма. Некоторые водные гифомицеты в условиях культуры на различных питательных средах образуют совершенную сумчатую стадию, относящуюся к родам нектрия (Nectria), моллизия (Mollisia), массарина (Massarina) и гименосцифус (Hymenoscyphus). У некоторых видов грибов из родов клавариопсис (Clavariopsis aquatica), трикладий (Tricladium gracile), варикоспорий (Varicosporium giganteum) известны дополнительные несовершенные стадии. Водные гифомицеты в культуре могут образовывать склероции. Особенностью поведения водных гифомицетов в культуре служит образование большинством из них на твердых питательных средах только неспороносящего мицелия. Образование конидий наблюдается в том случае, если кусочек мицелия вместе с агаром питательной среды перенести в сосуд со стерильной водой.
Водные гифомицеты играют немаловажную роль в самоочищении водоемов от растительных остатков, в первую очередь от листового опада, который ежегодно в обилии поступает в реки, ручьи, озера, пруды и водоемы других типов. Исследования по физиологии водных гифомицетов показывают, что все вещества, необходимые для роста этих грибов в качестве источников углерода и азота, они получают из погруженных в воду листьев, используя растительные белки и аминокислоты. Кроме того, водные гифомицеты в основном витамино-автотрофы, что также дает им определенные преимущества по сравнению с водными грибами других систематических групп при заселении и разложении органических остатков.
Известна роль водных грибов в качестве индикаторов разных типов и степеней загрязнения воды в водоемах. Водные гифомицеты из родов лунулоспора (Lunulospora curvula), лемонниера (Lemonniera aquatica), трикладий (Tricladium anomalum), клаватоспора (Clavatospora longibrachiata) и других служат показателями загрязнения водоемов отходами крахмало-паточного производства.
Водный гифомицет из рода центроспора (Centrospora acerina), обычно развивающийся как сапрофит на погруженных в воду листьях, вызывает гниль моркови, петрушки и пастернака в овощехранилищах, причем патогенными являются не только штаммы, выделенные из почвы или пораженных корнеплодов, но и из гниющих в воде листьев.
Сравнительно недавно открытая группа водных гифомицетов, несмотря на усиленное внимание к ней исследователей, все же во многих аспектах изучена еще недостаточно. Поэтому в ряде вопросов, связанных, в частности, с формированием этой группы, приходится говорить предположительно. Нет сомнения, что дальнейшие углубленные исследования водных гифомицетов раскроют интереснейшие моменты морфологии, биологии и экологии этой группы.
|
ПОИСК:
|
При использовании материалов проекта активная ссылка обязательна:
http://gribochek.su/ 'Библиотека о грибах'