Порядок Пероноспоровые (Peronosporales) (Н. С. Новотельнова)
Пероноспоровые по сравнению с сапролегниевыми и лептомитовыми представляют собой следующий этап приспособления грибов к жизни на суше. Это довольно древняя группа, возникновение которой относят к палеозойской эре (карбон), наиболее эволюционировавшая среди оомицетов. В ней насчитывается несколько сотен видов, причем наибольшее их число входит в семейство пероноспоровых (Peronosporaceae), представители которого характеризуются более совершенными признаками, тогда как другие семейства содержат значительно меньше видов и эволюционно менее прогрессировали.
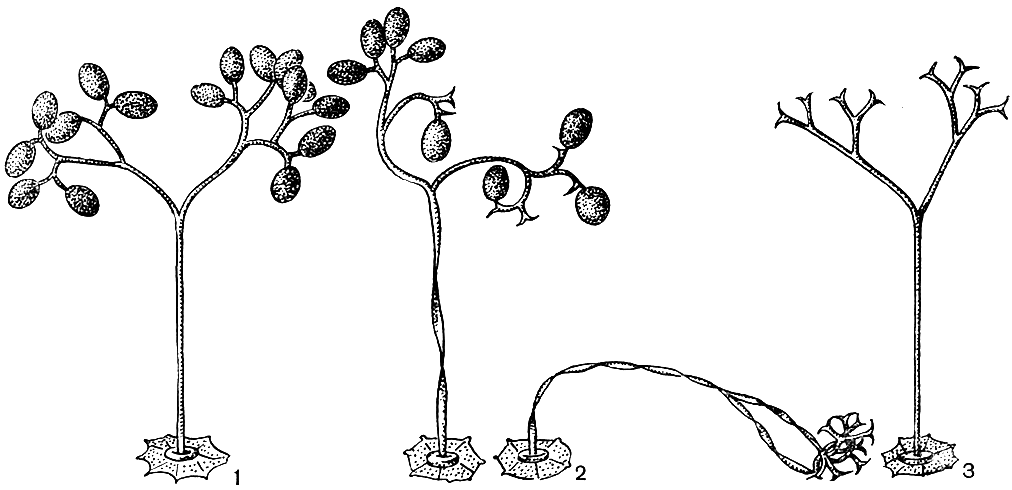
Рис. 22. Механизм разбрасывания конидий спороносцами пероноспоры (Peronospora tabacina): 1 - спороносец, выступающий из устьица; 2 - торзионное движение спороносцев; 3 - спороносец после сбрасывания спор
Порядок пероноспоровых делят на 3 семейства: питиевые, цистоповые и пероноспоровые.
По приуроченности к среде обитания виды пероноспоровых грибов можно отнести к гидрофилам, зоофилам и фитофилам.
Гидрофилами и зоофилами являются лишь некоторые представители семейства питиевых, тогда как преобладающее большинство видов относится к фитофилам. Соответственно этому у них выработались и различные приспособления для расселения в природе по воздуху, по воде, с помощью животных и человека.
Приспособления для распространения в водной среде сохранились у представителей всех семейств порядка. В цикле развития многих видов имеется стадия подвижных зооспор. Однако водная среда, играющая основную роль в распространении настоящих водных грибов, имеет у пероноспоровых грибов лишь ограниченное значение, обеспечивая только кратковременные связи гриба с водным субстратом.
Представителям родов питиум (Pythium), фитофтора (Phytophthora), склероспора (Sclerospora), базидиофора (Basidiophora), плазмопара (Plasmopara), цистопус (Cystopus) и отчасти пероноспора (Peronospora) свойствен особый механизм образования и выталкивания зооспор из их вместилищ - зооспорангиев, которые или отчленяются от спороносцев, или (у наиболее примитивных форм) остаются прикрепленными к несущим их гифам. Типично наземные представители семейства пероноспоровых выработали другие приспособления для распространения в природе. У них известно наличие особых гигроскопических торзионных механизмов. Древовидно разветвленные спороносцы под влиянием даже небольшого изменения влажности воздуха способны к вращению вокруг длинной оси. Вращение сопровождается сбрасыванием созревших спор с ветвей, и последние, подхваченные токами воздуха, переносятся на большие расстояния. Предполагается, что клеточные стенки спороносцев, состоящие у пероноспоровых грибов из целлюлозы, обладают спиральной структурой, и это определяет их торзионные свойства (рис. 22).
Расселение пероноспоровых грибов в природе происходит и при помощи человека. Этим способом в основном распространяются виды, паразитирующие на хозяйственно-ценных, интродуцированных человеком растениях. Клубни, плоды, луковицы, отводки, отдельные органы и целые растения, пораженные этими грибами, разносят своих спутников на дальние поля, в другие страны и на новые континенты.
Не уничтоженные и не обезвреженные остатки таких растений обеспечивают сохранение покоящихся форм пероноспоровых грибов в условиях почвы и возобновление видов из года в год.
Пероноспоровые грибы чрезвычайно широко распространены на земном шаре. Некоторые из них встречаются решительно повсюду, где произрастают высшие растения. К ним относятся, например, цистоповые грибы, обнаруживаемые и за полярным кругом, и в тропиках. При продвижении с юга на север количество видов заметно уменьшается. Благодаря особенностям своего развития и образу жизни наибольшее распространение они имеют в низменностях, где поселяются на равнинах в стациях, примыкающих к рекам, озерам и другим естественным и искусственным водоемам, отличающихся особым микроклиматом, главным образом высокой влажностью. Многие виды обитают в предгорных зонах, отдельные встречаются и в горах, преимущественно в сырых ущельях на теневыносливых растениях. В нижнем горном поясе, примерно до 1000 м над уровнем моря, способны обитать грибы довольно большого числа видов. В высокогорьях - в средней, верхней и субальпийской зонах - их заметно меньше. Но иногда Basidiophora entospora и отдельные виды родов Plasmopara и Peronospora поднимаются до высоты 3000-3800 м над уровнем моря, в зону, граничащую с вечными снегами. Определенные равнинные виды не поднимаются в высокогорья, а высокогорные виды не спускаются в равнины. Только высоко в горах встречаются, например, Peronospora dipelta и P. gypsophilae. В степях и пустынях также обитает незначительное число видов. Однако и в пустыне зарегистрированы отдельные виды пероноспоровых грибов. В пустынях Средней Азии встречается, например, Plasmopara pygmaea на анемоне, P. consolidae на дельфиниуме, в условиях пустыни центральной части Араратской долины иногда обильно развивается Peronospora effusa и другие виды.
По характеру развития пероноспоровые грибы - эфемеры. Они требуют короткого времени для образования спороношения и затем относительно быстро отмирают, но при помощи своих спор, попавших на благоприятные субстраты, развиваются заново, возрождаясь в новых поколениях, формирующих новые генерации спор, многократно повторяющиеся в течение вегетационного периода.
Наибольшего развития пероноспоровые грибы достигают в ранневесенний, весенний и весеннелетний периоды. В это время года, по существу, во всех зонах обитания нарастает численность видов и частота их появления. Некоторый подъем развития пероноспоровых грибов происходит и осенью, что связано с выпадением обильных осадков и смягчением температурного режима.
Самые ранние находки пероноспоровых грибов в средней полосе нашей страны относятся к апрелю - маю, когда они появляются на анемонах, сныти, дымянке, хохлатке. Преобладающее число видов отмечается в июне, наиболее благоприятном по количеству выпадаемых осадков. На севере в условиях Западной Сибири виды пероноспоровых грибов появляются позднее, обычно в последних числах мая; первыми из них под пологом осинового леса обнаруживаются на сныти Plasmopara aegopodii и на герани P. geranii-pratensis. Большинство видов развивается во второй половине июня, в первой и второй декадах июля, а в отдельные годы и в августе. В Средней Азии пероноспоровые грибы появляются значительно раньше. Даже в начале второй декады февраля можно увидеть Peronospora media, P. lepidiisativa, P. polygoni, однако столь же быстро, уже через месяц, развитие пероноспоровых грибов затухает, так как они не выносят низкую влажность воздуха и высокую температуру.
У пероноспоровых грибов тело состоит из грибницы, более или менее разветвленной, с гифами, довольно тонкими у представителей одних семейств (2-3 мкм) и относительно толстыми у других (7-12 мкм). В состав оболочек гиф входит целлюлоза. Мицелий, как правило, в начале развития несептированный, но впоследствии изредка образует перегородки, чаще всего при отделении старых участков гиф от нарастающих новых. Кроме того, на нем всегда возникают перегородки, вычленяющие органы спороношения.
Помимо однолетнего мицелия, у пероноспоровых грибов имеется мицелий многолетний, способный сохраняться в латентном состоянии внутри некоторых органов растений, например в луковицах, семенах и т. п., или продолжать активно развиваться в корнях, стеблях, стволах, ветвях многолетних растений.
У сапрофитных видов грибница стелется главным образом по поверхности субстрата, у паразитных приурочена к жизни в организме хозяина, разрастаясь там локально, в виде отдельного небольшого очага, или распространяясь диффузно по всему растению. Она пронизывает все его органы - корни, стебель, листья, цветки и цветоносы, плоды и плодоножки, семена (семенную оболочку), а иногда и завязь.
В процессе эволюции пероноспоровых грибов и адаптации к водным и наземным условиям существования у них выработались многие ценные приспособительные вегетативные признаки. Для длительного переживания при неблагоприятных условиях среды мицелий формирует хламидоспоры - образования с относительно толстой оболочкой и с большим запасом питательных веществ.
В оогонии возникает (в отличие от других оомицетов) лишь одна ооспора. Ооспоры, заполняющие (плеротические) оогоний, или ооспоры, не заполняющие (аплеротические) оогоний, у грибов разных видов имеют оболочку различной скульптуры - гладкую, складчатую, бугорчатую, бородавчатую, сетчатую, шиповатую (рис. 23). Известны случаи, когда ооспоры образуются без оплодотворения, партеногенетически.
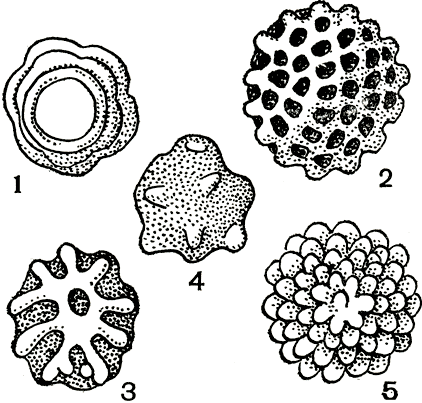
Рис. 23. Ооспоры пероноспоровых грибов: 1 - плазмопара на астрах (Plasmopara asterea); 2 - плазмопара на подсолнечнике (P. helianthi); 3 - пероноспора (Peronospora herniaris); 4 - цистопус (Cystopus tropicus); 5 - цистопус (С. candidus)
Своего рода "забота о потомстве" имеется у высших представителей порядка Peronosporales - членов семейств пероноспоровых и цистоповых. У этих облигатных паразитов оплодотворение по месту заложения половых органов обязательно внутреннее, глубинное, при котором зигота оказывается защищенной многими слоями клеток хозяина, тогда как у полупаразитов из семейства питиевых оплодотворение преимущественно поверхностное, наружное, не всегда защищенное покровами питающего растения. Количество закладываемых органов полового размножения обычно очень большое, в результате чего образуется неисчислимое множество ооспор, например у Sclerospora graminicola в листьях могара, S. secalina в листьях ржи, Plasmopara viticola в листьях винограда.
Но у отдельных представителей этого порядка ооспоры образуются не всегда, в небольшом количестве или совсем отсутствуют. Значение ооспор не только в постоянно происходящих процессах видо- и формообразования, но и для сохранения видов в природе. Ооспоры не менее года пребывают в латентном состоянии в остатках пораженных растений, попавших в почву, и часто не активизируются до полного разложения остатков, после чего под влиянием чередования намачивания, высушивания, нагрева и охлаждения они созревают, выходят из состояния покоя и становятся способны к ферментативной деятельности. Они прорастают затем с образованием ростковой трубки или зооспорангия. Этот процесс прослежен у паразитирующего на винограде вида Plasmopara viticola, но у многих видов еще не установлен.
Семейство Питиевые (Pythiaceae) (Ю. Т. Дьяков)
Питиевые грибы занимают промежуточное положение между водными сапрофитами из семейства сапролегниевых и высокоспециализированными паразитами высших наземных растений из семейства пероноспоровых. Это семейство очень важно для понимания хода эволюции у грибов. На его примере можно наблюдать, каким образом грибы расстались с водной средой и перешли к наземному образу жизни. Поэтому рассмотрение функциональной морфологии этих грибов, их физиологии и экологии представляет большой интерес.
Будучи переходным, это семейство, включающее всего несколько родов, необычайно широко распространено. Питиевые грибы встречаются в морской и пресной воде, в различных почвах и на растениях. Они найдены почти во всех точках земного шара - от Исландии до Южной Австралии, включая тропические зоны Африки, Азии и Америки.
Питиевые - одна из самых представительных групп почвенных грибов. Однако исследовать их истинную распространенность в почве не просто. Они медленно растут на обычных питательных средах и легко вытесняются быстро растущими несовершенными грибами, выделяющими к тому же в окружающую среду антибиотические вещества. Поэтому, если развести почвенный образец в стерильной воде и высеять на обычные для выращивания грибов питательные среды, питиевые грибы обнаружить не удастся.
Для выделения питиевых обычно применяют различные модификации двух методов. Первый метод заключается в добавлении в питательную среду антибиотиков, слабо действующих на питиевые, но подавляющих рост других почвенных грибов (например, полимиксина, пимарицина). Второй метод заключается в использовании ловушек, в которые избирательно переходят питиевые грибы. Такими ловушками могут служить простерилизованные с поверхности плоды различных растений (яблони, груши, огурца), наполовину погруженные во влажные почвенные образцы. Если на искусственных питательных средах питиевые растут медленнее большинства почвенных сапрофитов и вытесняются ими, то на живых растениях или их частях скорость распространения питиевых в связи с их паразитическими свойствами довольно высока. Помещая загнившие выше поверхности почвы части плодов на питательные среды, можно выделить из них питиевые грибы многих видов. Таким образом удалось получить объективную оценку истинного распространения питиевых грибов в почвах, причем по числу изолятов они оказались на одном из первых мест среди почвенных грибов.
Образ жизни питиевых грибов так же разнообразен, как и места их обитания.
Обитающие в водной среде грибы питаются главным образом водорослями. Среди них встречаются как сапрофитные, так и паразитические формы. Питиевые грибы паразитируют на многих видах зеленых водорослей (из класса равножгутиковых, сцеплянок и харовых), красных (Porphyra, Ceramium), сине-зеленых (Tolypothrix). Эти паразиты малоспециализированы. Они могут развиваться в клетках водорослей из различных классов и даже отделов, а при искусственном заражении поражают и высшие растения. Например, Pythium adherens паразитирует на водорослях Rhizoclonium hierogliphicum, Spirogyra crassa, Ulothrix zonatae, Tolypothrix sp. и может заражать кукурузу, горох, помидоры, огурцы. Некоторые водные питиевые грибы встречаются на живых и мертвых беспозвоночных животных (например, на дафнии).
При паразитировании на водорослях мицелий большинства питиевых грибов развивается непосредственно в клетках хозяев, он пронизывает эти клетки и вызывает их гибель (рис. 24). Некоторые виды, однако, воздействуют на свою жертву иначе. Они образуют выросты мицелия - гаустории, которые проникают в клетку хозяина и служат питательным органом. Пораженные клетки дольше остаются живыми, а это выгодно паразиту.
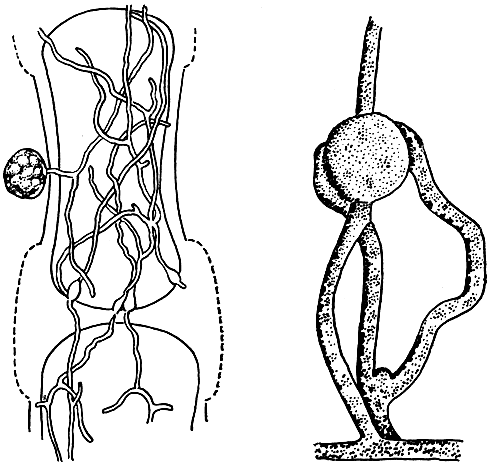
Рис. 24 (слева). Поражение красной водоросли церамиум питиумом (Pythium maritimum): 1 - мицелий внутри клеток водоросли; 2 - освобождение зооспор
Почвообитающие питиевые грибы, так же как и водные, ведут сапрофитный или паразитический образ жизни. Большинство из них поражают подземные органы высших растений. Более примитивные виды могут неопределенно долгое время находиться в почве в сапрофитном состоянии, питаясь растительными остатками, но при наличии в почве корневой системы растений способны ее поражать. При этом питиевые поражают чаще всего молодые растения, корни которых не покрыты еще хорошо развитыми защитными покровами, а также растения, ослабленные неблагоприятными условиями роста. Пораженные корешки отмирают, а надземные части растения отстают в росте и часто также погибают.
Поражение корней питиевыми грибами носит название корнееда или корневой гнили. Корневой гнилью поражаются всходы сахарной свеклы, хлопчатника, люцерны и деревьев (рис. 29,1). Особенно сильно корнеед развивается в годы с холодной и влажной весной, когда корневая система развивается медленно, а отдельные участки корней вследствие недостатка воздуха в переувлажненной почве отмирают. Мертвые клетки не способны к активной физиологической защите и легко заселяются грибами, служа воротами инфекции. Поселившись на отмерших участках корней, паразит питается там и выделяет токсические вещества, которые убивают прилегающие к мертвым живые участки. Таким образом осуществляется продвижение паразита по ткани хозяина. Подобные виды питиевых грибов, так же как и паразиты водорослей, широко специализированы и могут заражать десятки и сотни видов высших растений из разных родов и семейств.
У других видов питиевых паразитические свойства еще более развиты. В отсутствие растений-хозяев они могут находиться в почве лишь в покоящемся состоянии, ибо вегетативный мицелий не выдерживает конкуренции с сапрофитными почвенными грибами и быстро погибает. Некоторые из них поражают не только корневую систему, но и надземные части растений. Это высокоспециализированные паразиты высших растений. Для заражения такие грибы уже не нуждаются в участках отмерших тканей, а питаются лишь живыми клетками хозяина. Поэтому их мицелий развивается внутри тканей по межклетникам, а в клетках находятся лишь выросты мицелия - гаустории, служащие для питания. Зараженные клетки долго остаются жизнеспособными.
Циклы развития и морфологические особенности питиевых грибов отражают образ их жизни. Вегетативный мицелий одет целлюлозной оболочкой, содержит много ядер и способен к неограниченному росту в питательном субстрате, будь то почва или ткани растения-хозяина. На мицелии образуются органы бесполого и полового размножения. Первые представлены зооспорангиями, в которых развиваются зооспоры. Зооспорангии водных и многих обитающих в почве питиевых представляют собой концы недифференцированных гиф, отделенные от остального мицелия перегородкой. У одних грибов (Pythium dissotocum, P. monospermum) зооспорангии, как и у сапролегниевых, нитевидные; у других (P. debaryanum и фитофтора) зооспорангии округлые или лимоновидные. Паразитические питиевые грибы, мицелий которых развивается внутри тканей хозяина, для распространения зооспор должны иметь особые выросты мицелия - зооспорангиеносцы, выносящие зооспорангии на поверхность субстрата.
Способ прорастания зооспорангиев зависит прежде всего от образа жизни. У всех водных и большинства почвенных грибов оболочка на вершине или сбоку зооспорангия разрывается и его содержимое выходит наружу в виде пузыря, в котором протоплазма разделяется на отдельные участки, превращающиеся в зооспоры. Оболочка зооспорангия при этом остается на мицелии. Такой способ прорастания связан с тем, что выходящие из зооспорангия зооспоры сразу попадают в благоприятные для них условия - в водную среду, в которой они могут активно двигаться и прорастать. У видов, поражающих надземные части растений, зооспорангии формируются на поверхности растений, т. е. в условиях быстрой смены увлажнения и высыхания. Зооспоры, представляющие собой комочки протоплазмы без оболочек, не способны даже короткое время находиться вне капель воды. Поэтому при описанном выше способе прорастания был бы большой риск гибели вышедших из зооспорангиев зооспор в связи с подсыханием листьев.
Прорастание зооспорангиев у подобных видов разделено на два этапа. Сначала зооспорангии в целом виде отделяются от спорангиеносцев. Они имеют плотные оболочки и способны сохранять жизнеспособность и вне капель воды. Зооспорангии разносятся ветром и дождевыми брызгами. Поэтому они способствуют распространению гриба в пространстве (у водных и почвообитающих видов эту функцию несут активно плавающие зооспоры). Выход зооспор из зооспорангиев возможен только в каплях воды и при пониженной температуре, т. е. в условиях, гарантирующих медленное высыхание капель. Вышедшие из зооспорангиев зооспоры движутся непродолжительное время (от нескольких минут до нескольких часов) и прорастают. Если зооспорангии проросли в каплях на поверхности восприимчивых растений, ростки зооспор внедряются в растительные ткани, в иных случаях проросшие зооспоры погибают.
При повышенной температуре (больше 20° С) капли могут быстро высыхать, поэтому у некоторых видов питиевых грибов в процессе эволюции возникла способность к прорастанию отделившихся от зооспорангиеносцев зооспорангиев в таких условиях непосредственно ростковой трубкой, которая внедряется в растительные ткани. Таким образом, в семействе питиевых в зависимости от условий обитания можно наблюдать все варианты бесполого размножения, от типичных зооспорангиев, характерных для водорослей и водных грибов, до конидий, характерных для наземных высших грибов.
После выхода из зооспорангия зооспоры претерпевают три этапа - движение, инцистирование и прорастание.
Движение обеспечивают два жгутика. Обычно зооспоры пассивно движутся с током воды. Однако у многих видов, паразитирующих на корнях высших растений (Pythium aphanidermatum, Phytophthora fragariae, P. parasitica и др.), обнаружено активное движение в направлении корней. Особенно сильно зооспоры привлекаются пораненными участками корней (рис. 26). Известно, что именно такие участки служат воротами инфекции, поэтому движение в их направлении позволяет зооспорам эффективнее осуществлять заражение своих хозяев.
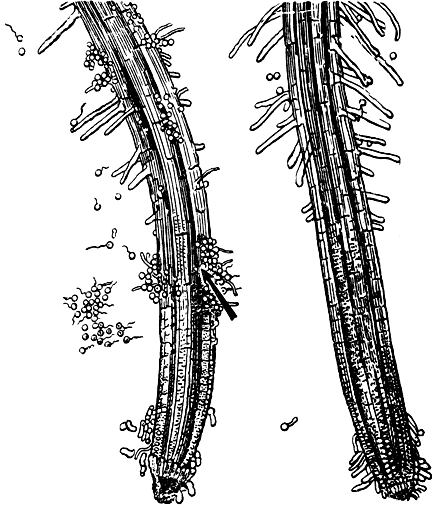
Рис. 26. Привлекающее действие пораненных участков корней для зооспор питиума (Pythium aphanidermatum). Стрелкой указано пораненное место. Справа - непораженный кончик корня
Удалось установить причины привлекающего действия корней. Например, зооспоры P. palmivora активно двигаются в направлении корней кокосовой пальмы. В экстрактах кожуры найден ряд аминокислот, которые, как и корни, привлекают паразита. Поскольку при поранении выделение химических веществ в окружающую среду усиливается, становится понятной причина привлечения зооспор к пораненным участкам корня.
Помимо химических, зооспоры питиевых грибов могут привлечь и физические факторы. Когда зооспоры P. parasitica помещали в камеры с электродами, наблюдали скопление зооспор у катода. Если электроды заменяли корешками, зооспоры скапливались у корешков. Поскольку корни выделяют слабые токи, возможно, движение по направлению к корням обусловлено электротаксисом, который обнаружен у нескольких видов питиевых грибов.
Значение таксисов для этих грибов, однако, не очень велико. Без помощи токов воды зооспоры могут пройти лишь очень ограниченное расстояние. Например, скорость хемотаксического движения зооспор Pythium aphanidermatum составляет около 2,5 мм в минуту. В увлажненной почве, помещенной в чашки Петри, зооспоры Phytophthora cinnamomi достигали кончиков корней, удаленных от них не больше 7 см.
После периода движения зооспоры теряют жгутики, округляются и одеваются тонкой оболочкой - инцистируются. Вслед за инцистированием начинается прорастание зооспор ростковой трубочкой.
Органы полового размножения образованы выростами мицелия. Женские органы (оогонии) - шарообразные выросты, а мужские (антеридии) - нитевидные (рис. 25). Кончик антеридия прикладывается к оогонию, их оболочки в месте соприкосновения растворяются, и содержимое антеридия переливается в оогоний. После слияния двух ядер образуется диплоидная ооспора. Она имеет толстую оболочку, много питательных веществ и способна долго сохраняться в неблагоприятных условиях. У некоторых видов питиума установлено, что ооспоры сохраняют жизнеспособность в воздушно-сухой почве больше 12 лет. Прорастают ооспоры зооспорангием или ростковой трубкой.
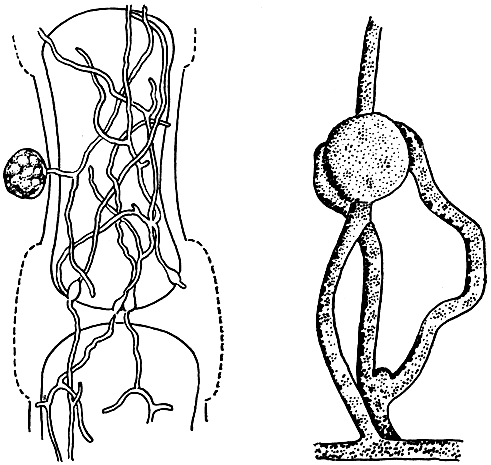
Рис. 25 (справа). Оогонии и антеридии питиума (Pythium fabae)
Многие питиевые грибы гомоталличны, т. е. на одном и том же мицелии образуются как антеридии, так и оогонии. Однако встречаются и гетероталличные виды, у которых для образования половых органов необходим контакт мицелиев разных штаммов. У гетероталличных видов выделяют два изолята - А1 и А2. Ооспоры образуются лишь при совместном росте изолятов А1 и А2. В месте контакта мицелиев кончик гифы одного изолята закручивается и утолщается, давая начало образованию антеридия. Контактирующая с антеридием гифа другого изолята начинает прорастать через него, после чего над антеридием образуется сферическое тело - оогоний. Антеридий у таких видов имеет форму воротничка, окружающего шейку оогония.
Изоляты разных типов спаривания А1 и А2 нельзя назвать мужским и женским. Один и тот же штамм А1 при встрече с одними штаммами А2 образует антеридии, т. е. обладает мужским полом, а при встрече с другими может образовывать оогонии, становясь изолятом женского пола. Следовательно, гетероталличные питиевые грибы потенциально обоеполы, их раздельнополость относительна. Контакт гиф разных типов спаривания лишь стимулирует мицелии к образованию половых органов. Если между мицелиями грибов, относящихся к разным типам спаривания, поместить целлофан, предотвращающий непосредственный контакт между гифами, но позволяющий обмениваться продуктами обмена веществ, гетероталличные изоляты могут превратиться в гомоталличные, т. е. на одном и том же мицелии возникнут антеридии и оогонии, а в дальнейшем и зрелые ооспоры. Следовательно, толчок к образованию половых органов у гетероталличных видов дают химические вещества, проходящие через целлофан.
В семействе питиевых различают несколько родов.
Представители рода зоофагус (Zoophagus) имеют нитевидные зооспорангии, отделенные от несущей их гифы перегородкой. Они паразитируют на коловратках и имеют на мицелии специальные короткие разветвления, приспособленные для захватывания коловраток (рис. 27, 1). В свободном виде они встречаются в воде между нитями водорослей. У грибов из рода питиогетон (Pythiogeton) зооспоры формируются в виде недифференцированной массы в длинном пузыре, выходящем из зооспорангия, а ооспоры одеты очень толстой оболочкой. Развиваются они главным образом на погруженных в воду ветках и плодах (рис. 27, 2).
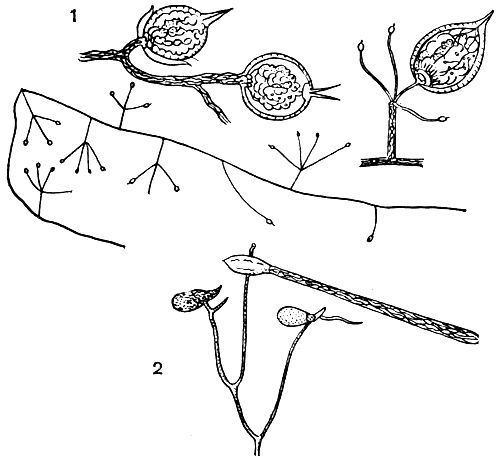
Рис. 27. 1 - мицелий зоофагуса (Zoophagus tentaclum) с ветвями, пронизывающими коловраток; 2 - спороношение питиогетона (Pythiogeton transversum)
Благодаря видовому богатству, широте распространения и практическому значению наибольший интерес представляют грибы из родов питиум (Pythium) и фитофтора (Phytophthora).
Представители рода питиум имеют тонкий, паутинистый мицелий (толщина гиф 3-6 мкм), простирающийся по пищевому субстрату. Большинство водных питиумов обитает в пресной воде, однако Pythium marinum, P. maritimum (рис. 24) и P. reptans встречаются на живых и мертвых морских красных водорослях Сеramium, Porphyra, Bangia.
Pythium marinum живет как паразит и сапрофит морских красных водорослей Ceramium, Porphyra. Мицелий этого гриба заражает клетки водорослей с помощью присосок - аппрессориев - и тянется через много клеток. Вытянутые спорангии пробивают клеточную стенку хозяина и протягиваются на короткие расстояния в воде. Зооспоры образуются в пузыре, выходящем из зооспорангия. Заражение разных видов водорослей ростовыми трубками зооспор происходит по-разному. При заражении водоросли Ceramium гриб образует присоски и тонкую "инфекционную" гифу, развивающую большое давление и, как шило, пробивающую клеточную стенку водоросли. С помощью таких инфекционных структур происходит прорыв оболочки и внедрение паразита в клетку. Оболочка водоросли Porphyra, вероятно, прободается легче, поэтому инфекционных структур в данном случае не образуется.
Большинство пресноводных грибов рода питиум паразитирует на зеленых нитчатых водорослях. Наиболее распространены морфологически близкие виды P. gracile и P. tenue. У первого из них оогонии крупнее, чем у второго (16-20 мкм против 7-17 мкм), и одеты более толстой оболочкой.
Антеридии и оогонии у P. gracile всегда образуются на разных гифах, в то время как у P. tenue они могут быть и на одной гифе. Оба вида встречаются в клетках спирогиры, где они образуют разветвленный мицелий с обильными половыми органами и ооспорами (рис. 28).
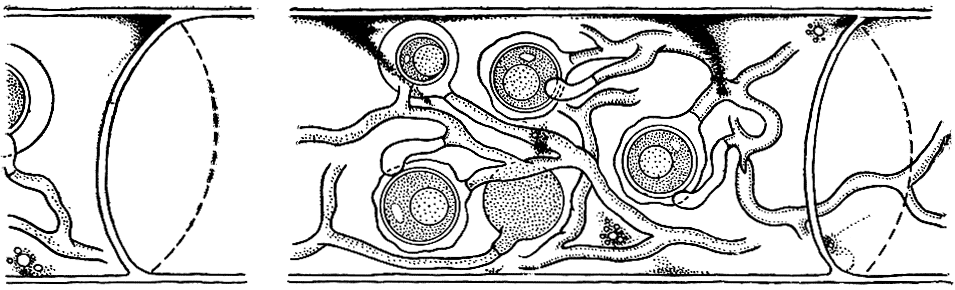
Рис. 28. Мицелий и половые органы питиума (Pythium gracile) в клетках зеленой водоросли спирогиры
Кроме спирогиры, грибы из рода Pythium паразитируют на водорослях Rhizoclonium, Cladophora, Vaucheria, Sphaeroplaea, Chara, Nitella, Chlorococcum и др.
Некоторые виды (например, P. daphnidarum) поражают мелких ракообразных.
Обитающие в почве питиумы поражают корни наземных растений, вызывая корневые гнили. Несколько видов (например, P. marschancia) выделено из мхов, остальные заражают семенные растения.
Из 133 видов питиума около 40 найдено в водной среде, остальные - в почве.
В почве встречается большинство из известных видов этого рода. Круг растений-хозяев, образ жизни и морфология почвообитающих видов очень схожи, поэтому точно определить видовую принадлежность выделенных грибов не всегда удается.
Pythium debaryanum вызывает корневую гниль всходов многих растений. Он описан в качестве паразита на сахарной свекле, огурцах, лесных породах и других растениях в СССР, Западной Европе и Америке. В нашей стране этот гриб особенно вредит сахарной свекле. Поражается преимущественно подземная часть всходов сахарной свеклы. Пораженные ткани буреют и загнивают, растения привядают, семядоли и первые листочки желтеют. Сильное поражение происходит перед "линькой" коры, когда первичная кора отмирает и теряет свои защитные функции. После образования вторичной коры (в фазу двух пар настоящих листьев) всходы становятся устойчивыми к заражению. В пораженных тканях образуются органы бесполого и полового размножения гриба (рис. 29).
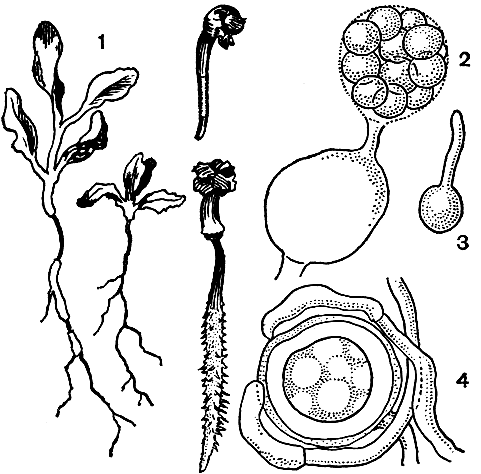
Рис. 29. Питиум (Pythium debaryanum): 1 - поражение проростков и всходов сахарной свеклы корневой гнилью; 2 - выход зооспор из зооспорангия в пузыре; 3 - проросшая зооспора; 4 - оогоний с двумя антеридиями
Долгое время P. debaryanum считался самым распространенным видом этого рода. При более тщательном изучении морфологии питиевых грибов оказалось, однако, что несколько морфологически близких видов встречаются в почве и пораженных растениях чаще, чем P. debaryanum.
Pythium irregulare (табл. 7,2) по внешнему виду и паразитической специализации близок к P. debaryanum, отличаясь от него главным образом наличием выростов оболочки оогония (протуберанцев). Особенно вредит этот гриб всходам древесных пород. Например, он чаще других видов питиума выделяется из лесных питомников США и Австралии; из корней пораженных персиковых деревьев в Америке; в СССР выделен из корневой системы сахарной свеклы, которую, в отличие от P. debaryanum, может поражать в течение всего вегетационного периода, вызывая черный сосудистый некроз. У зараженных растений отмирают сосуды листьев и корней, поэтому растения отстают в росте от здоровых и часто загнивают.
P. ultimum имеет сильно разветвленный мицелий. Зооспор не образуется, и зооспорангии прорастают главным образом ростковой трубкой, гриб поражает всходы более 150 видов растений (на всех континентах), среди них - сахарную свеклу, огурцы и другие тыквенные, цитрусовые, лесные деревья. Распространению спор P. ultimum в почве способствуют нематоды - мелкие круглые черви, питающиеся на корнях растений. Есть сведения о том, что этот гриб может вступать в симбиотические отношения с корневой системой ряда растений, образуя микоризу.
P. aphanidermatum (табл. 7,3) поражает более 80 видов растений. Особенно сильный вред он наносит злаковым культурам (ржи, овсу), плодовым (яблоне, груше), огурцам, сое, томатам, сахарной свекле и другим растениям.
P. sylvaticum вызывает большой вред в лесных питомниках Европы и Америки, однако поражает также землянику, пшеницу, лен, горох и др. Этот вид гетероталличен, причем показано, что изоляты, на которых формируются оогонии, более патогенны, чем антеридиальные изоляты. Поскольку патогенность большинства питиевых грибов связана с наличием токсина, который выделяется в окружающую среду и убивает зараженные клетки, можно предположить, что оогониальные изоляты выделяют больше токсина, чем антеридиальные.
В роде фитофтора насчитывают около 70 видов. По морфологическим, физиологическим и экологическим особенностям этот род занимает промежуточное положение между питиевыми и пероноспоровыми грибами.
Мицелий грибов фитофторы белый, паутинистый. Спорангии обычно лимоновидной формы, зооспоры формируются внутри зооспорангия и выходят через разрыв оболочки, образующейся на вершине зооспорангия. В воде эти грибы не развиваются и могут попасть туда лишь случайно. Большинство из них, подобно почвообитающим питиевым, поражают подземные органы растений и образуют спороношение лишь во влажной почве; зооспоры распространяются почвенными водами. Однако, за редким исключением, грибы фитофторы не могут длительное время существовать в почве, где сохраняются лишь их покоящиеся органы - ооспоры и хламидоспоры. В отличие от питиевых они из почвы, как правило, не выделяются, но их можно легко выделить из пораженных частей растений. Некоторые виды, подобно пероноспоровым грибам, поражают надземные части растений.
Поскольку все фитофторовые грибы ведут активный паразитический образ жизни, у них возникли различные приспособления для паразитирования на высших растениях. На примере рода фитофтора можно совершить увлекательную экскурсию в прошлое и рассмотреть, как проходила эволюция грибов от водных к наземным, от сапрофитов к паразитам. Этот род уникальный в мире грибов в том отношении, что нигде больше морфологически близкие виды настолько сильно не отличаются по физиологии питания и образу жизни. В роде фитофтора длительный процесс эволюции паразитизма развернут как бы в одной плоскости.
Самые примитивные паразиты из этого рода могут длительное время существовать в почве, заселяя, подобно питиевым грибам, растительные остатки. Они заражают растения через раны, выделяют сильные токсины, разрушающие клетки и ткани хозяев. Поскольку они питаются уже частично разрушенными тканями и даже растительными остатками, им почти безразличен вид растения-хозяина; поэтому они поражают все, что попадается на пути.
Специализация их в отношении видов растений-хозяев очень широкая, как и у видов рода питиум. Однако, в отличие от последних, они характеризуются и широкой специализацией в отношении поражаемых органов и тканей. Грибы рода питиум в основном поражают лишь подземные органы, как бы боясь расстаться с привычной средой обитания - почвой. Фитофторовые приобрели способность поражать не только корни, но и листья, плоды, кору, камбий, т. е. практически любые органы и ткани. Однако мицелий их, развиваясь внутри тканей, прободает клетки хозяев и вызывает их гибель. Выделяемые этими грибами токсины разрушают окружающие клетки, поэтому зараженная ткань обычно загнивает, а спороношение гриба образуется на отмерших тканях, попадающих в почву, как и у представителей питиума.
Следовательно, гриб из почвы заражает растение-хозяина, быстро убивает его и возвращается в почву, совершая как бы отдельные набеги на соседние территории, но не задерживаясь там после их разграбления. Такие набеги вызваны стремлением уйти от конкуренции с многочисленными почвенными микроорганизмами. Ведь почва буквально насыщена сапрофитными микроорганизмами (в 1 г почвы обитают миллионы бактерий и актиномицетов, десятки и сотни тысяч зародышей грибов). Всем им нужны богатые органическим веществом растительные остатки; многие микроорганизмы выделяют антибиотики, губительно действующие на соседей.
Осваивая ткани и органы растений, недоступные в связи с наличием защитных свойств большинству почвенных сапрофитов, гриб приобретает для себя несомненные преимущества. Но для этого надо, чтобы его хозяин не погибал как можно дольше. На место кратковременных набегов с разграблением соседних территорий приходит длительная их колонизация. На примере рода Phytophthora можно наблюдать, как это происходит. У некоторых видов появляется способность к специализированному заражению живых тканей. Вместо ран воротами инфекции становятся устьица. Мицелий развивается между клетками, а для питания появляются специализированные выросты мицелия - гаустории. Пораженные клетки в результате этого дольше остаются живыми. Но за эти преимущества паразит расплачивается дорогой ценой. Чтобы не убивать своего хозяина, гриб неизбежно должен был потерять самые агрессивные орудия нападения - токсины и гидролитические ферменты, разрушающие растительные ткани. Находясь в среде, богатой готовыми органическими веществами, паразит частично утрачивает способность самостоятельно синтезировать их из более простых продуктов; не испытывая конкуренции со стороны других микроорганизмов, он разучился бороться с ними. Поэтому возврат в почву стал для таких паразитов невозможен, что еще более усилило зависимость их от растений-хозяев. Поскольку обмен веществ у неродственных растений различен, а паразит утратил свое ферментативное богатство, произошло сужение паразитической специализации до отдельных семейств и даже родов высших растений. Все эти переходы можно наблюдать у отдельных представителей фитофторы.
Таким образом, род фитофтора можно считать мостом, соединяющим примитивных паразитов из рода питиум с высокоспециализированными паразитами семейства пероноспоровых. В этом роде осуществлены почти все эволюционные завоевания морфологического и физиологического характера, сделавшие фитофтору одним из самых процветающих паразитов высших растений.
Фитофтора широко распространена по земному шару и встречается почти во всех климатических зонах. Наиболее богаты ее видами тропические и субтропические области, которые надо считать центрами возникновения рода. Для умеренного пояса наибольшее распространение и практическое значение имеют виды Phytophthora infestans и P. cactorum, для субтропиков - P. parasitica, для тропиков - P. palmivora и P. cinnamomi.
Пожалуй, нет среди грибов другого, столь трагически известного гриба, как P. infestans. Это единственный гриб, о котором есть сведения, в каком году он появился в каждой стране, в каком месяце и даже какого числа появилась болезнь, им вызываемая. Этот гриб оказал трагическое влияние на судьбу целой нации. Написанных о нем книг и статей достаточно, чтобы занять целую библиотеку. И сейчас, несмотря на то что его, казалось бы, обуздали, он время от времени дает о себе знать и вызывает опустошительные вспышки болезни, которую разные народы называли "картофельной гнилью", "картофельной чумой" и "холерой".
Картофель, как известно, был завезен в Европу из Америки. Вскоре в странах Северо-Западной Европы он стал основным продуктом питания бедных слоев населения. О том, как важен был картофель для бельгийских шахтеров, свидетельствует, например, известная картина Ван-Гога "Едоки картофеля". В Европе картофель ежегодно давал высокие стабильные урожаи. Одной из причин этого было отсутствие болезней, которые он оставил на своей родине - в Центральной и Южной Америке. Картофельная болезнь появилась почти одновременно в США и в Европе (1842-1843). Возможно, ее привезли из Перу в связи с массовым вывозом чилийской селитры, начавшимся в 1840 г., а в 1844 г. болезнь уже была зарегистрирована в Бельгии, Франции (Лилль), Англии (Кент и Суссекс) и Шотландии. Но сухое лето не способствовало ее распространению. 1845-1846 годы стали драматической вехой в судьбе народов Северо-Западной Европы.
В середине июля 1845 г. был поражен картофель на всей территории Фландрии и прилегающих областях Франции и Голландии. Августовские бельгийские газеты уже были заполнены статьями о новой болезни картофеля. Затем болезнь перешла в Люксембург и по долинам Роны и Рейна - в Швейцарию. В середине августа наблюдалось массовое поражение картофеля в окрестностях Парижа. Положение складывалось настолько серьезным, что члены Французской Академии наук были отозваны из отпусков.
В середине августа болезнь распространилась в районы нижнего Рейна и Южной Англии. Первое сообщение о картофельной гнили в Ирландии было опубликовано в дублинской вечерней газете 6 сентября. Вначале там не придавали большого значения болезни, но при уборке урожая в октябре выяснилось, что большинство клубней гнилые. Это вызвало панику.
В том же году болезнь распространилась на восток - в Пруссию, Польшу и Скандинавию, однако потери здесь вследствие сухой осени были невелики.
В следующем году болезнь появилась сначала в Ирландии и распространялась там со скоростью 50 миль в неделю на восток и северо-восток. Уже в начале августа большинство посевов погибло.
На территории России первые сообщения о картофельной гнили появились в Прибалтике, где в 1846-1847 гг. болезнь вызвала гибель большинства посевов картофеля. Затем она распространилась в Белоруссию и Северо-Западную Россию.
Картофельная болезнь принесла в Европу голод и нищету. Особенно сильно она отразилась на судьбе Ирландии. В 1845 г. население Ирландии составляло около 8 млн. человек. Из них для 6 млн. картофель занимал по крайней мере половину пищевого рациона, а остальные питались почти исключительно картофелем. Лишившись его, люди лишились единственного источника существования. Очевидцы описывают селения, жители которых походили на обтянутые кожей скелеты; у них не было даже сил подниматься с постелей. Некоторые селения полностью вымерли. Вид гниющих растений вызывал у населения ужас. Бедствие усугублялось тем, что люди боялись употреблять в пищу пораженные клубни. За голодом последовали его неминуемые спутники - инфекционные болезни. Началась массовая эмиграция оставшихся в живых ирландцев в Англию и США. К 1851 г. население Ирландии уменьшилось на 2 млн. человек. "Что значит золотуха,- писал Ф. Энгельс,- в сравнении с тем голодом, который постиг в результате болезни картофеля Ирландию и который свел в могилу миллион питающихся исключительно или почти исключительно картофелем ирландцев, а два миллиона заставил эмигрировать за океан!" (К. Маркс и Ф. Энгельс. Соч., т. 7, с. 12). К. Маркс в статье "От февраля до июня 1848 года" оценил вспышку картофельной болезни как "событие мирового значения" (К. Маркс и Ф. Энгельс. Соч., т. 20, с. 497).
Загадку таинственной картофельной болезни разрешил А. де Бари, установивший инфекционную природу заболевания, описавший возбудителя и цикл его развития. Название, которое дал де Бари возбудителю, в переводе на русский язык означает "пожиратель растений инфекционный".
Возбудитель картофельной болезни - гриб Phytophthora infestans (рис. 30, табл. 6) - развивает внутри листьев картофеля межклеточную грибницу с гаусториями. Питаясь тканями листа, он вызывает образование темных пятен, которые во влажную погоду чернеют и загнивают. При сильном поражении отмирает весь лист (табл. 6). Сначала погибают нижние листья, а затем болезнь захватывает и все растение. После периода питания на грибнице образуются выросты - спорангиеносцы, которые высовываются наружу через устьица (табл. 6,4). Во влажную погоду они образуют налет белого цвета вокруг пятен с нижней стороны листьев (табл. 6,2). На концах спорангиеносцев формируются лимоновидные зооспорангии, которые отрываются и разносятся ветром или брызгами дождя. Попадая в капли воды на поверхности листа картофеля, спорангии прорастают шестью - восемью зооспорами, которые после периода движения округляются, покрываются оболочкой и прорастают ростковой трубкой (рис. 30, 1-4). Росток через устьице проникает в ткань листа (рис. 30, 5). При благоприятных условиях (прохладная дождливая погода или сильные росы) время от заражения до образования нового спороношения составляет всего 3-4 дня. Так как вокруг одного пятна образуются тысячи зооспорангиев, можно понять, насколько быстро распространяется болезнь в посевах картофеля.

Таблица 6. Фитофтороз картофеля и томатов: 1 - поражение листьев картофеля; 2 - поражение плодов томатов; 3 - поражение клубней картофеля
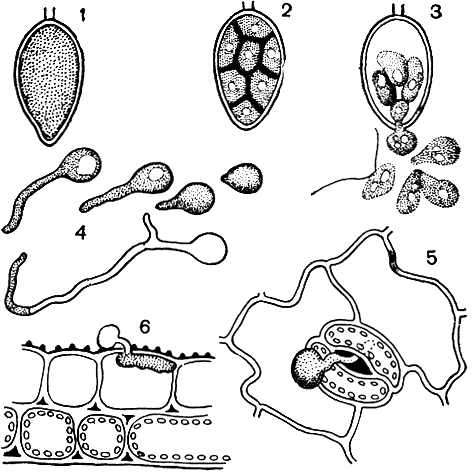
Рис. 30. Возбудитель картофельной болезни - фитофтора (Phytophtora infestans): 1-3 - стадии прорастания зооспорангия; 4 - прорастание зооспор; 5 - внедрение через устьице; 6 - внедрение в ткань листа через кутикулу
Попадая на землю, спорангии профильтровываются через верхние слои почвы и вызывают заражение формирующихся клубней. Массовое заражение клубней происходит во время уборки, при соприкосновении пораженной ботвы с пораненными участками клубней. На клубнях болезнь проявляется в виде свинцово-серых пятен. Ткань клубня под пятном буреет, но не размягчается (табл. 6, 2). Иногда побурение захватывает значительную часть клубня. Такие клубни во время хранения сгнивают. Пораженные клубни подвержены сильному заражению различными бактериями и грибами, что усиливает вредоносность фитофтороза.
Инфекция "охраняется в слабо пораженных клубнях, при высадке которых в поле на ростках образуется спороношение гриба. Однако долгое время гриб почти не размножается, и лишь в июле - августе наблюдается массовое развитие болезни. В почве спорангии и мицелий долго сохраняться не могут и быстро погибают, поэтому почвенная инфекция не играет большой роли.
P. infestans - гетероталличный гриб, т. е. для образования половых органов необходимо слияние изолятов А1 и А2. В Европе, однако, гриб развивается только в бесполой стадии и ооспор не образует. Половой процесс обнаружен лишь на его родине - в Мексике, где найдены оба типа спаривания. Все европейские и североамериканские изоляты относятся к одному типу - А1, что обусловливает невозможность образования ооспор. Почему, несмотря на интенсивный обмен, изоляты типа А2 не попали до сих пор из Мексики даже в США, остается загадкой. Вероятно, эти изоляты неоднократно попадали в другие зоны, но условия не способствовали их существованию. Гриб сформировался в Мексике, природные условия которой лучше всего благоприятствуют его развитию. В горных районах Мексики есть долина Толука, которую П. М. Жуковский назвал "санаторием для фитофторы".
В Толуке длинные (12 ч) холодные и влажные летние ночи. Во второй половине дня почти ежедневно идут ливневые дожди. Зимы сухие и теплые. Ооспоры гриба могут там сохраняться в течение всей зимы в почве. Прорастающие ооспоры заражают всходы картофеля на уровне почвы. Массовое поражение наблюдается уже через 40 дней после посадки картофеля (в Европе - не раньше, чем через 90-100 дней). Поэтому в Толуке создана научная станция по изучению фитофтороза картофеля.
Кроме картофеля, гриб поражает томаты и некоторые сорные пасленовые. Поражение томатов может быть не менее сильным, чем картофеля. Гриб поражает листья и плоды, на которых возникают бурые пятна, делающие плоды непригодными в пищу (табл. 6,5). Сильный вред томатам болезнь приносит на Дальнем Востоке, где муссонные дожди способствуют ее развитию, а также на северо-западе СССР и в районах поливного овощеводства Украины и Закавказья.
Борьба с фитофторозом картофеля и томатов ведется в двух направлениях: химической защитой и выведением устойчивых сортов.
В годы первой вспышки картофельной болезни (в 1845-1846 гг.) газетный корреспондент из Уэльса сообщил, что картофельной гнили нет на полях, расположенных вблизи медеплавильных заводов, между тем как на полях, удаленных от медеплавилен, картофель превратился в вонючую гниющую массу. Однако прошло еще много лет, пока опрыскивание ботвы картофеля смесью медного купороса с известью (бордоской жидкостью) стало общепринятым. Помимо бордоской жидкости, сейчас в борьбе с фитофторозом картофеля употребляют много новых фунгицидов (веществ, убивающих грибы). Химическая обработка всегда уменьшает зараженность и гарантирует получение удовлетворительного урожая даже в благоприятные для развития болезни годы. Но это повышает трудоемкость выращивания картофеля, удорожает его и вызывает накопление в почве и клубнях остатков вредных химических веществ.
Другой путь защиты от болезни - селекция устойчивых сортов картофеля. Ее история начинается с экспедиций Н. И. Вавилова в Центральную и Южную Америку за дикими видами картофеля. Мексика оказалась уникальным источником устойчивых к фитофторозу видов. Здесь на совместной родине хозяина и паразита сформировались высокоиммунные виды, скрещивание которых с культурным картофелем привело к созданию устойчивых сортов.
Казалось, проблема картофельной гнили, столь долго волновавшая мир, решена. Но через несколько лет начали поступать сообщения о том, что новые сорта тоже стали поражаться болезнью. Начались тщательные исследования причин потери устойчивости, приведшие к обнаружению высокой лабильности гриба, популяции которого не однородны, а состоят из различных по вирулентности рас.
Выращивание устойчивых сортов привело к тому, что расы, способные их заражать, быстро размножились и вызвали массовое их поражение. Снова селекционеры принялись за работу и вывели сорта, устойчивые к более вирулентным расам. Увы! Успех опять оказался лишь временным. Такое соревнование между человеком и грибом продолжается до сих пор. Борьба с грибом оказалась более сложной, чем предполагалось раньше. Но уже появились новые идеи в селекции, новые фунгициды. Давно ушли в безвозвратное прошлое трагические эпифитотии середины XIX в. Несомненно, в будущем вредоносность болезни будет сведена до минимума.
Гриб Phytophthora cactorum (табл. 7) впервые был описан как паразит декоративных кактусов в Германии в 1875 г. Существует мнение, что он перешел на декоративные растения американского происхождения от сапрофитного образа жизни или раньше поражал сорные растения в Европе. В 1904 г. в Швейцарии было обнаружено поражение P. cactorum яблонь и груш, а в 1912 г.- земляники. В 1922 г. гриб был найден в Югославии, в 1933 г.- в Швеции, в 1934 г.- в Дании и в 1938 г.- в США. В СССР поражение земляники было впервые обнаружено в 1950 г., а яблони - в 1957 г.

Таблица 7. Пероноспоровые грибы: 1 - фитофтороз женьшеня: пораженные корень и листья; 2 - питиум нерегулярный (Pythium irregulare) на корнях растений гороха; 3 - питиум афанидерматум (P. aphanidermatum) на соприкасающихся с почвой плодах тыквы; 4 - базидиофтороз: налет гриба на листе мелколепестника; 5 - склероспора злаковая (Sclerospora graminicola) на могаре: разрушеннные грибом листья; 6 - плазмопара подсолнечниковая (Plasmopara helianthi): карликовое растение с налетом гриба на нижней стороне листьев; 7 - плазмопара белоснежная (Р. nivea) на сныти: налет гриба на листьях
В отличие от высокоспециализированного паразита, каким является P. infestans, этот гриб находится на самом низком уровне паразитической специализации. P. cactorum - раневой паразит, образующий сильные токсины. Его мицелий проходит внутри тканей хозяина как межклетно, так и через клетки, не образуя гаусторий. Гриб очень широко специализирован, поражая 83 рода высших растений из 44 семейств, однако все же отмечена наибольшая приуроченность его к поражению розоцветных (поражается 13 видов), бобовых (11 видов) и сосен (10 видов). Злаки совершенно не поражаются. Особой приуроченности к поражению каких-либо органов гриб не имеет, вызывая корневые гнили всходов (особенно у сосен), гнили корней (у бобовых), гнили основания стебля (у лилий), гнили коры и плодов (у розоцветных), гнили сердцевины и почек (у рододендронов, пионов).
Наибольшее практическое значение имеют поражения грибом плодовых и ягодных растений из семейства розоцветных в умеренной зоне. На землянике фитофтора вызывает кожистую гниль, при которой плоды становятся бурыми и горькими. Пораженная ткань ссыхается, поверхность становится кожистой и покрывается густым белым налетом. На яблонях гриб вызывает раковые трещины коры у основания ствола, гниль корней, листьев и плодов.
P. parasitica - также очень широко специализированный гриб, поражающий растения 72 родов из 42 семейств. Но, в отличие от Р. сасtorum, он образует межклетный мицелий с гаусториями, т. е. стоит на более высоком уровне паразитической специализации.
P. parasitica (рис. 31) - наиболее широко распространенный представитель рода фитофтора, но самый большой ущерб он вызывает в субтропических странах. Несмотря на широкую специализацию, замечается некоторая приуроченность гриба к пасленовым растениям (отмечен на восьми видах пасленовых в 40 странах). Наибольшее экономическое значение имеют вызываемые этим паразитом гнили корней и стеблей у томатов, табака и перцев. Кроме того, гриб может вызывать также сильные поражения тыквенных растений, хлопчатника, цитрусовых, папайи и др. В умеренной зоне он может причинять большой урон культуре томатов в оранжереях. Ооспоры и хламидоспоры этого гриба сохраняются в почве.
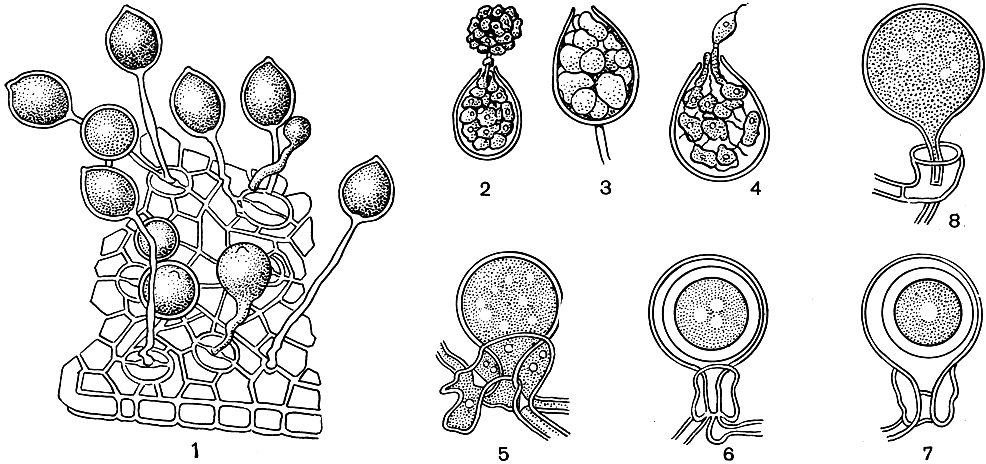
Рис. 31. Фитофтора (Phytophtora parasitica): 1 - спорангиеносцы со спорангиями, выходящими через устьица; 2-4 - прорастание зооспорангиев; 5-8 - стадии образования ооспор
P. palmivora - один из самых распространенных паразитов тропических растений. Он поражает 51 род из 29 семейств высших растений, вызывая гнили корней и плодов папайи и хлебного дерева на островах Тихого океана, гниль какао в Бразилии, Мексике, Гватемале, Нигерии и на Ямайке, гниль гевеи на Шри Ланка, в Малайе, Бразилии и Коста-Рике, гниль пальм в Африке и Юго-Восточной Азии, гниль цветков берберы в Индии, гниль перца в Южной Америке и Юго-Восточной Азии. Особенно страдают от этого гриба кокосовые пальмы. Он поражает их листовые влагалища, которые буреют, загнивают и покрываются во влажную погоду налетом спороношения гриба. Очень сильно поражается папайя на Гавайских островах. Паразит вызывает гниль корней папайи. В сезон дождей обильный налет зооспорангиев и хламидоспор покрывает нижнюю часть ствола и плоды, которые, падая на землю, вносят в почву инфекцию. Оттуда зооспоры могут попасть на молодые корни соседних растений и заразить их.
Другой способ распространения спор гриба - не через почву, а по воздуху. Было обнаружено, что плодовые мушки дрозофилы охотнее питаются загнившими плодами папайи, нежели здоровыми. На своем теле эти насекомые распространяют спорангии гриба.
Несмотря на широкую специализацию, P. palmivora стоит на более высокой степени паразитической специализации, чем P. cactorum. У P. palmivora мицелий развивается межклетно с гаусториями. Гриб гораздо слабее связан с почвой и способен распространяться по воздуху, да и широта специализации, возможно, кажущаяся. Есть много данных о том, что P. palmivora - сложный вид, состоящий из морфологически одинаковых штаммов, которые, однако, специализированы к различным видам хозяев. Это такой момент в процессе эволюции паразитизма, когда распадение широкоспециализированного вида на отдельные узкоспециализированные формы не привело еще к появлению морфологических различий между ними, т. е. специализированные формы не обособились еще до ранга отдельных видов.
Разделение P. palmivora по физиологической специализации было впервые сделано американским микологом Эшби, который в 1922 г. выделил 2 группы - с гевеи и какао. Изоляты с какао лучше заражали какао, чем гевею, а изоляты с гевеи, наоборот, лучше заражали гевею. После того как у P. palmivora был установлен гетероталлизм, нашли, что штаммы с гевеи чаще относятся к группе А1, а штаммы с какао - к группе А2.
В дальнейшем оказалось, что специализацией обладают и изоляты с других видов растений. Например, изоляты, выделенные из перца, при искусственном заражении могли заражать лишь разные виды перца. Ни один из 43 испытанных видов растений, относящихся к 20 семействам, не заразился. Это свидетельствует о наличии узкоспециализированных групп внутри широкоспециализированного вида.
P. cinnamomi - один из самых активных грибных паразитов растений. Впервые был описан в качестве паразита корицы и авокадо на Суматре, однако в настоящее время с помощью человека распространился по всему земному шару. Гриб поражает 212 видов, относящихся к 117 родам из 48 семейств папоротников, голосеменных и покрытосеменных растений. Большинство его хозяев - тропические и субтропические растения (авокадо, корица - рис. 32, орхидеи и др.), но, попав в более умеренные зоны, гриб вызвал разрушительные болезни местных растений, такие, как корневая гниль всходов сосны в США и Новой Зеландии, чернильная болезнь каштана (поражение камбия) в Европе, гниль грецкого ореха в США и Европе, гниль рододендрона, азалии, вереска и других растений во многих частях света. В СССР P. cinnamomi обнаружена в Грузии, где она поражает пробковый дуб, грецкий орех и хинное дерево. Растения некоторых видов иммунны к P. cinnamomi. Например, сеянцы цитрусовых за один год устраняют инфекцию из сильно зараженной почвы. Этот гриб имеет некоторые свойства, сближающие его с сапрофитными почвенными грибами, т. е. находится на невысокой ступени паразитической специализации. В отличие от большинства видов фитофторы он может некоторое время развиваться в почве, ведя там сапрофитный образ жизни. Во влажной почве при ее естественном заражении гриб без своих растений-хозяев может сохраняться больше шести лет, а в воздушно-сухой почве погибает через 3 месяца. В почве сохраняются мицелий и хламидоспоры. Ооспоры образуются лишь на пораженных корнях. Органы бесполого размножения - зооспорангии - возникают редко. Для их образования необходимо стимулирующее воздействие выделений некоторых почвенных микроорганизмов, поэтому в стерильных условиях на искусственных средах они почти никогда не образуются.
Семейство Пероноспоровые (Реronosporасеае) (Н. С. Новотельнова)
Это наиболее крупное семейство порядка пероноспоровых, содержащее наиболее высокоорганизованных представителей. По своим чертам семейство значительно отличается от предыдущих. Все виды - наземные формы, поселяющиеся исключительно на живых растениях. Все они - облигатные паразиты, поражающие при благоприятных условиях растения в любом возрасте и активно воздействующие на своего хозяина в течение вегетационного периода. В остатках пораженных растений, находясь в латентном состоянии, они доживают до следующего вегетационного периода и таким образом возобновляются из года в год.
Связи пероноспоровых с питиевыми можно усмотреть не только в морфологической близости, но в некоторых биологических особенностях видов, например в способности (правда, немногих представителей) заселять корни растений (Plasmopara helianthi, Peronospora ficariae). Сохранившаяся в цикле развития отдельных видов стадия зооспор также свидетельствует об этих связях. Большинство же грибов семейства пероноспоровых заселяют листья растений в ряде случаев цветки (Peronospora corollae, P. radii).
Семейство представлено пятью родами - Basidiophora, Sclerospora, Plasmopara, Peronospora, Bremia. Основные особенности строения органов бесполого размножения те же, что и у других пероноспоровых,- отсутствие особых приспособлений для защиты от вредного воздействия факторов среды (нагрева, охлаждения, инсоляции и т. п.). Роды в структурном отношении четко различаются. Общая для всех черта (за исключением Basidiophora) - древовидная форма спороносцев. Структурные признаки указывают на наличие двух основных направлений морфогенеза этих грибов в процессе эволюции, обусловивших родовое разнообразие. Одно из них - это усложнение ветвления спорангиеносцев от простого и беспорядочного к более сложному и четко выраженному (Sclerospora, Plasmopara, Peronospora); другое - появление новой, особой структуры спороносца с расширенной вершиной (неразветвленного у Basidiospora и разветвленного у Bremia). Первое направление морфогенеза возникло, несомненно, ранее второго, что можно заключить по его более примитивным показателям и приуроченности видов к паразитированию на цветковых растениях таких семейств, как лилейные, злаковые, лютиковые и др.; второе - значительно позже, что явствует из специализации видов Basidiophora и Bremia в пределах наиболее высокоорганизованного из цветковых растений семейства сложноцветных.
Грибы семейства пероноспоровых появляются ранней весной на семядольных листьях еще слабых всходов растений, а затем на настоящих листьях, цветках, плодах, реже на стеблях и корнях, вызывая иногда местное поражение, но чаще болезнь всего растения. Присутствие гриба обнаруживается в виде светло-зеленых, угловатых или расплывчатых, постепенно буреющих пятен на верхней стороне листовых пластинок и войлочного налета белого, серого, серовато-фиолетового или серовато-ржавого цвета на нижней. При диффузном поражении растения гриб сильно угнетает процессы его роста. Больное растение превращается в карлика, малооблиствленного и неплодоносящего; оно недолго вегетирует и обычно вскоре засыхает. Распространение грибов и заражение ими растений происходит в течение всего вегетационного периода, однако жаркая и сухая погода не только задерживает, но и исключает развитие паразитов. Повышенная влажность воздуха и умеренная температура способствуют нашествию пероноспоровых грибов на растения в полях, лугах, лесах и зачастую ведет к значительному преобладанию их над другими видами паразитных грибов.
Для мицелия грибов семейства пероноспоровых характерно наличие гаусторий (пузыревидных, пальцевидных, простых и разветвленных), проникающих в клетки растения-хозяина. Как правило, межклеточный мицелий не выходит из тканей в воздушную среду, а образующийся поверхностный налет - это бесполое спороношение гриба, состоящее из густого сплетения спороносцев, несущих споры, осуществляющие расселение вида и заражение новых растений. Половое спороношение возникает внутри тканей во всех органах хозяина и выявляется часто уже весной. Однако это больше связано с наступлением жаркого летнего периода, а осенью - холодного. Роль ооспор в жизненном цикле видов не всегда ясна, так как прорастание ооспор у отдельных видов, по наблюдениям исследователей, сомнительно. В этих случаях гриб резервируется, по-видимому, в зимующих органах растений, в корнях многолетников или в семенах.
Благодаря приспособительным структурным и биологическим признакам пероноспоровые грибы относительно легко адаптируются к условиям среды и многие их виды очень широко расселены в природе, таковы Plasmopara viticola, P. helianthi, Peronospora tabacina, P. destructor.
Хотя все виды пероноспоровых - облигатные паразиты, у каждого проявляется своеобразие во взаимоотношениях с питающим его организмом. Так, например, нападение одних грибов на растения осуществляется через корневые волоски; других - только через устьица (рис. 33); наконец, третьих - через непосредственное прободение эпидермальных клеток. Эти грибы приурочены к определенным органам и тканям, а развитие их зависит от условий среды.
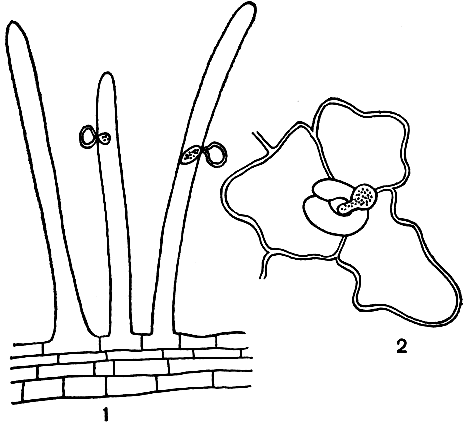
Рис. 33. Внедрение ростков зооспор в растения: 1 - зооспоры плазмопары (Plasmopara helianthi), проникающие в корневой волосок подсолнечника: 2 - зооспора плазмопары (P. viticola), проникающая в ткань листа винограда через устьице
Некоторые роды пероноспоровых приурочены к определенным семействам цветковых растений. Например, только Sclerospora поражает злаки, только Plasmopara - зонтичные и виноградовые, только Peronospora - бобовые, крестоцветные, розоцветные и др. Но на некоторых семействах цветковых паразитируют различные роды пероноспоровых, хотя не на одном и том же виде растения-хозяина, а на разных видах. Например, на лютиковых и гераниевых встречаются Plasmopara и Peronospora, на сложноцветных - Basidiophora, Plasmopara, Peronospora и Bremia.
Некоторые пероноспоровые грибы, поражающие цветковые растения одного и того же семейства, довольно узкоспециализированы в пределах рода, а иногда и вида питающего растения, но морфологически мало отличаются друг от друга. При тщательной ревизии они зачастую оказываются не видами, а специализированными формами, или расами. В других случаях их можно считать самостоятельными видами по комплексу свойств (ареал, физиологические особенности) даже при слабо выраженных морфологических различиях. Это относится к виду Peronospora tabacina, видам Plasmopara на зонтичных, видам Peronospora на крестоцветных, на бобовых и т. д.
Род базидиофора (Basidiophora) (табл. 7) относится к наиболее редко встречающимся пероноспоровым грибам, хотя и распространен во многих странах Европы, Америки, Австралии. Он необычен по своему строению и не имеет аналогов ни в порядке пероноспоровых, ни вообще среди низших грибов. По типу спороношения у него обнаруживается структурное сродство с некоторыми высшими грибами (рис. 34,2). Характерные для рода спороносцы не разветвлены, на вершине булавовидно вздуты и по форме напоминают базидии гименомицетов.
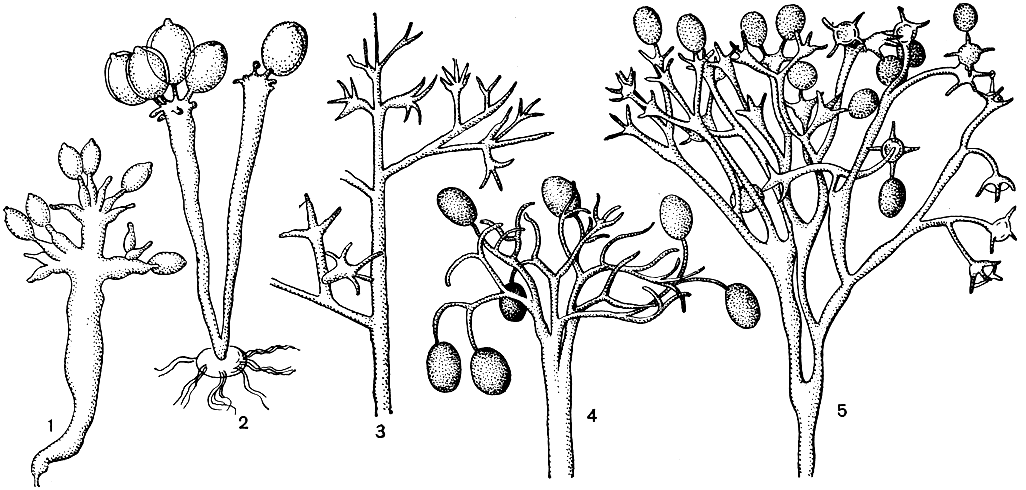
Рис. 34. Конидиеносцы пероноспоровых грибов: 1 - склероспора; 2 - базидиофора; 3 - плазмопара; 4 - пероноспора; 5 - бремия
Род представлен всего двумя видами - В. entospora и В. kellermannii. Первый из них поражает прикорневые листья мелколепестника (табл. 7,4). Спороносцы выступают на поверхность листьев из устьиц небольшими группами, образуя нежный беловатый налет. На их вершинах на небольших коротких отрогах одиночно возникают лимоновидные споры. К периоду созревания споры отчленяются с небольшой "ножкой" - частью несущего отрога. Споры являются типичными зооспорангиями и, помещенные в воду, прорастают с образованием зооспор, которые выходят наружу через вершинный сосочек. Одновременно внутри тканей листьев на межклеточном ветвистом мицелии, снабженном пузыревидными гаусториями, возникают органы полового размножения - оогонии и антеридии, а затем формируются желтоватые, с толстой, многослойной, складчатой оболочкой ооспоры. Этот вид был впервые найден во Франции в 1868 г., а впоследствии обнаруживался повсеместно, но нечасто в различных климатических зонах. В. entospora поражает также золотарник и астры, а В. kellermannii - циклахену дурнишниколистную.
Род склероспора (Sclerospora) распространен более широко и содержит около 20 видов. Представители его встречаются в Европе, Азии, Америке и Африке, причем большинство видов приурочены к тропическим и субтропическим областям.
Грибы этого рода поражают лишь растения из семейства злаковых, в том числе такие важнейшие сельскохозяйственные культуры, как пшеница, рожь, ячмень, кукуруза, просо, сорго, могар, сахарный тростник. Грибы вызывают пятна на листьях, образуя с нижней стороны пластинки обычный для пероноспоровых грибов налет, размочаливание листа и гипертрофию разных органов.
Мицелий склероспоры межклеточный, ветвистый, иногда с пузыревидными гаусториями. Спороносцы низкорослые, с беспорядочно расположенными толстыми ветвями, на концах с небольшими отрогами, несущими одиночные споры без сосочка (рис. 34,1). У грибов многих видов это спороношение не развивается (S. secalina) или настолько эфемерно, что его редко удается заметить. Вообще бесполое спороношение у отдельных видов не играет существенной роли в их жизненном цикле. Преимущественное значение у них имеет половая стадия развития. Ооспоры образуются внутри тканей хозяина в таком множестве, что становятся заметными даже при рассмотрении пораженных листьев без лупы.
К наиболее известным и распространенным видам относится S. graminicola (табл. 7), паразитирующий на растениях щетинника, сорго и проса. На листьях проса этот гриб вызывает беловатый, матовый, нестирающийся налет, не изменяя общего облика растения. В то же время паразит нещадно истребляет растения могара. В зараженном растении он развивается очень быстро и заполняет все его органы. К завершению цикла развития гриба все ткани листа, кроме проводящих, оказываются "нафаршированными" ооспорами. Эти участки листьев обезвоживаются, высыхают, разрушаются и вываливаются из листа. При этом ооспоры высыпаются в виде золотистой пыли. От листьев остаются мочаловидные пучки (табл. 7,5). Нарушается и внешний вид растений, которые становятся кустистыми, образуют многочисленные пасынки и мелкие, уродливые и бесплодные метелки.
В СССР S. graminicola распространена на могаре в Омской области и на Алтае, а на щетиннике зеленом повсеместно.
S. secalina относится к видам ограниченно распространенным, однако встречается не только на севере, где был вначале обнаружен Н. А. Наумовым (Архангельская область), но и на юге (Львовская область), а недавно найден в Прибалтике. Особенностью этого вида являются его сокращенный цикл развития и способность к образованию одной лишь стадии ооспор в листьях растений, на которых он паразитирует. Гриб поражает лишь рожь Secale cereale. По сравнению с другими видами особо сильного воздействия на своего хозяина он не оказывает.
Вредоносны S. maydis и S. philippinensis, паразитирующие на кукурузе, S. sacchari, S. spontanea - на сахарном тростнике и других злаках.
Род плазмопара (Plasmopara) содержит несколько десятков видов, из которых многие широко распространены на разных континентах земного шара. Род характеризуется спорангиеносцами, многократно моноподиально разветвленными, как правило, под прямым углом, на вершинах с 3-4 небольшими отрогами, несущими споры (рис. 34,5). Последние овальные, часто бочковидные, с сосочком на вершине и ножкой. Прорастание осуществляется образованием зооспор.
Гриб сохраняется в природе и возобновляется из года в год при помощи ооспор или зимующего мицелия, который снабжен пузыревидными гаусториями.
Плазмопары паразитируют на цветковых растениях многих семейств, вызывая тяжело протекающие заболевания, заканчивающиеся гибелью растений. Они поражают преимущественно травянистые растения, изредка кустарники. Заболевание проявляется диффузно, с поражением всего растения и сопровождается отставанием роста, карликовостью, или местно, в виде угловатых, просветленных, как бы маслянистых, растекающихся вдоль жилок пятен на листьях, с белым, сероватым или буроватым налетом.
Многие представители плазмопары (грибы разнообразных видов, разновидностей, подвидов и форм ее) паразитируют на растениях семейства лютиковых - ветреницах, пролесках, василистниках, живокости.
На крыжовниковых изредка встречается P. ribicola, на гераниевых паразитируют P. pusilla, P. geranii-pratensis, P. geranii-sylvatici; на бальзаминовых - P. obducens; на виноградовых - P. viticola, P. amurensis, P. chinensis.
Многие представители семейства зонтичных подвергаются нападению грибов из рода плазмопара: P. aegopodii паразитирует на сныти (табл. 7), P. angelicae - на дуднике, P. chaerophylli - на пупыре, P. laserpitii - на гладыше, P. sii - на поручейнике, а другие виды - на борщевике, горичнике, будренце, гирче, подлеснике, любистоке, болиголове, моркови, сельдерее, пастернаке.
Некоторые грибы рода плазмопара, паразитирующие на хозяйственноценных культурах, вызвали катастрофические бедствия в тех странах, в которых они впервые появились. В прошлом веке в Европу из Северной Америки был завезен возбудитель милъдъю винограда (P. viticola), нанесший неисчислимый ущерб виноградникам Франции, а потом и других стран, особенно в средиземноморской зоне. Распространился он также и в Азии и в Африке. Гриб поселяется на виноградной лозе, поражая листья, усики, цветки, плодоножки, ягоды. Его развитие зависит от температуры и влажности. В благоприятные для него годы с обильными дождями и росами он способен уничтожить значительную долю урожая винограда, накопляясь в природе сначала на порослях и одичавших кустах винограда.
В. Мюллер (1922) установил корреляцию между инкубационным периодом развития паразита и температурой. Эта зависимость, показанная для условий Бадена (ФРГ), выражена автором в виде кривой, получившей название "Кривая Мюллера". Позднее к ней были сделаны дополнения для других зон виноградарства, в том числе в нашей стране (H. И. Олтаржевский, Д. Д. Вердеревский). P. viticola поражает также другие виды винограда.
Не менее вредоносен возбудитель ложной мучнистой росы подсолнечника P. helianthi (табл. 7), завезенный в Европу из Северной Америки в XX в. Он распространен во многих странах Европы, Азии и Америки почти во всех зонах производственного возделывания подсолнечника. Нападая на это растение, гриб поражает все его органы и, поселяясь в точке роста, воздействует столь сильно на ростовые процессы, что такое мощное в здоровом состоянии растение превращается в слабого карлика (табл. 7,6). Его деформированные листья покрыты белым налетом, крошечная корзинка, потерявшая свойство гелиотропизма, тесно прижата к стеблю и несет пустые или неполноценные семена. Гриб способен значительно снизить в благоприятные для него годы урожай семян подсолнечника. P. helianthi содержит морфологические и специализированные формы, приуроченные к разным видам подсолнечника; ему свойствен необычный для пероноспоровых грибов полиморфизм.
Из видов плазмопары, поражающих культурные растения, менее вредоносны и не ежегодно встречаются виды, паразитирующие на моркови, фенхеле, сельдерее, петрушке, пастернаке, а также на астрах и золотарнике.
Род бремия (Bremia) своеобразен в семействе пероноспоровых, отличительные признаки которого достаточно выделяют его среди остальных родов семейства. По особенностям строения спороносного аппарата и образу жизни бремия считается одним из наиболее продвинутых в эволюционном отношении и полностью наземным. Относительно высокое положение видов бремия в порядке пероноспоровых подтверждается также тем, что эти грибы обнаруживаются избирательно на растениях наиболее высокоорганизованного семейства сложноцветных. Характер паразитирования, наличие рас, приуроченных к растениям одного только рода и даже вида питающего растения, - также свидетельство значительного эволюционного совершенства грибов рода бремия.
Отличительный морфологический признак рода - древовидно разветвленные конидиеносцы, с ветвями дважды и трижды дихотомически разветвленными, имеющими на вершинах дланевидные расширения с сидящими на них 2-8 короткими отрогами с тупыми концами. Каждый отрог несет по одной конидии (рис. 34,5). Последние снабжены на вершине сосочком, через который, однако, выходят не зооспоры, а росток мицелия. Внутри тканей питающего растения в межклетниках образуются оогонии и антеридии, там же возникают в результате полового процесса зимующие споры с гладкой или слегка бородавчатой оболочкой желтовато-коричневатого цвета.
Бремия вызывает сильное поражение растений. Грибы проникают в листья обычно непосредственно через эпидермис и развивают в тканях хозяина межклеточный мицелий с мешковидными гаусториями, вызывая диффузное или местное поражение с типичными для представителей семейства симптомами.
Виды бремии паразитируют на растениях семейства сложноцветных: В. centaurea - на васильках, В. lampsanae - на бородавнике и осоте, В. tulasnei - на крестовнике и т. д. Наиболее известен и широко распространен во многих странах мира вид В. lactucae, паразитирующий на салате (табл.8). Этой культуре гриб в отдельные годы наносит существенный ущерб. В составе его имеются многочисленные специализированные формы, поражающие дикие и сорные растения (свыше 125 видов), в том числе поповник, лопух, одуванчик (табл. 8,5), осот, бодяк, девясил и др.

Таблица 8. Пероноспоровые грибы: 1 - пероноспора чистяковая (Peronospora ficariae): налет на листьях ветреницы; 2 - пероноспора разрушающая (P. destructor): пораженный цветонос лука; 3 - пероноспора хмеля (Pseudoperonospora humuli): колосовидный побег, налет на листьях; 4 - цистопус кандидус (Cystopus candidus): искривленный стебель, видоизмененные стручочки, пустулы на листьях пастушьей сумки; 5 - бремия салатовая (Bremia lactucae): налет гриба на листьях одуванчика
Род пероноспора (Peronospora) наиболее богато представлен видами в семействе пероноспоровых и всего порядка Peronosporales. Он насчитывает около 300 видов, из которых большинство широко распространены в природе.
Виды пероноспоры поражают только травянистые растения из многих семейств. Мицелий у них с гаусториями разного типа. Аппарат бесполого размножения по своему характеру тот же, как и у остальных родов. Спорангиеносцы дихотомически разветвленные, с конечными ветвями прямыми или согнутыми, короткими или относительно длинными, раздвоенными под прямым или острым углом, заостренными на вершине, несущими одиночные споры (рис. 34,4). Последние овальные, чаще эллипсоидальные, без сосочка и ножки. Оогонии и антеридии образуются внутри тканей растения; ооспоры шаровидные, с гладкой, складчатой, бородавчатой или сетчатой оболочкой, желтовато-коричневые.
Род пероноспора содержит 2 подрода, различающихся по характеру прорастания спор бесполого размножения. Подрод псевдопероноспора (Pseudoperonospora) морфологически идентичен с родом Peronospora, но по биологическим свойствам он более примитивен. Споры здесь настоящие зооспорангии и прорастают с образованием зооспор. Подрод эупероноспора (Euperonospora) включает виды, утерявшие в процессе эволюции функцию формирования зооспор. Их споры - типичные конидии без вершинного сосочка, дающие при прорастании мицелиальные ростки, выходящие наружу непосредственно через оболочку в любой части ее поверхности.
Все грибы рода пероноспора - вредоносные паразиты, вызывающие болезнь и гибель растений. Симптомы поражения местные и общие - пятна, налеты, деформации, карликовость. Эти грибы не вызывают поражения корней, они преимущественно поражают листья как всходов, так и взрослых растений. Некоторые виды приурочены к репродуктивным органам растений. Вид Peronospora corollae, обнаруженный В. Г. Траншелем на венчиках колокольчика и льнянки, растет бессимптомно в зараженных весной проростках, следуя за точкой роста до момента цветения, и только тогда сиороносит на лепестках венчика. Аналогично ведет себя гриб P. radii на венчиках цветков различных сложноцветных - тысячелистника, пупавки, ромашки, а P. tranzscheliana - на цветках норичниковых, P. violaceae - на цветках ворсянковых, P. Scutellariae и P. stigmaticola - на цветках губоцветных.
Виды пероноспоры широко распространены на растениях семейства лютиковых - лютике, ветренице (табл. 8,1), чистяке, живокости; гвоздичных - звездчатке, ясколке, дрёме, горичнике; крестоцветных - бурачке, котране; розоцветных - лапчатке, черноголовнике. Известно много видов этого рода, паразитирующих на растениях семейства бобовых - пажитнике, астрагале, остролистнике, вике, чине, сое, люцерне, клевере; на лилейных, норичниковых, маковых, тыквенных, крапивоцветных, коноплевых, гречишных, маревых, молочайных, дымянковых, первоцветных, вьюнковых, пасленовых, бурачниковых, губоцветных и т. д. Из них целый ряд видов - опасные для культурных растений паразиты.
Pseudoperonospora humuli (табл. 8) из представителей подрода псевдопероноспоры, паразитирующий на диком хмеле и культурных сортах хмеля, распространен в Европе, Азии и Америке. Поражая молодые побеги, гриб вызывает их "колосовидность" - укорочение междоузлий, ненормальное обилие мелких, скрученных, морщинистых листьев. Уродуются главные и вторичные побеги, появляются так называемые белые побеги, частично лишенные хлорофилла. Гриб поселяется также на листве, казалосьбы, здоровых побегов и вызывает их засыхание и отмирание с образованием желтовато-зеленоватых пятен неправильной, угловатой формы. Позднее гриб переходит на цветки, а затем и на шишки хмеля, которые остаются недоразвитыми и бледно-зелеными, а затем буреют, засыхают и преждевременно опадают. На пораженных органах гриб образует довольно плотный войлочный серовато-фиолетовый до бурого налет, а внутри тканей - ооспоры (табл. 8, 5). Очень вредоносны виды P. cubensis для огурцов, дынь, тыкв, P. cannabina - для конопли.
Пероноспора табака (Peronospora tabacina) обнаружена впервые в 1850 г. в Австралии. Затем этот гриб распространился на всех континентах земного шара. На территорию Европы он проник в 1959 г., когда одна английская фирма по производству средств защиты растений завезла вид P. tabacina из США для своих опытов, не предприняв специальных мер безопасности. С ошеломляющей быстротой гриб в том же году расселился в странах Европы, в первую очередь в Голландии, Германии, а затем и в других европейских странах.
В течение вегетационного периода гриб образует многие генерации спор, хорошо переносит зимовку в климатических условиях табаководческой зоны и, ежегодно возобновляясь, наносит существенный ущерб растениям табака в теплицах и на плантациях. Он настолько вредоносен, что поставил под угрозу табаководство во многих странах мира.
Рассада заражается весной от перезимовавших в растительных остатках ооспор (источником инфекции могут быть и семена с пораженных растений). Сильно пораженные растения обычно гибнут, а другие, перенесенные в полевые условия, становятся центрами очагов распространения болезни.
Обычно на пораженных растениях табака появляются сначала одиночные, затем многочисленные желтоватые, округлые или угловатые, ограниченные жилками буреющие пятна на нижних, а потом на верхних листьях, края которых становятся волнистыми и заворачиваются книзу. Бурые пятна возникают и на жилках, черешках, стебле, корне. При влажной и дождливой погоде на нижней стороне пораженных листьев возникает спороношение гриба; оно голубоватого цвета (отсюда название болезни "голубая плесень"), но постепенно становится сероватым.
Заболевание скоротечно, особенно при поражении проростков, которые сначала увядают, а затем засыхают и отмирают. Более старые растения иногда оправляются. Поселяясь на листьях растения, гриб делает их негодными для промышленного использования. Гриб проникает в цветки и поражает образующиеся семена.
На пасленовых паразитирует также и P. hyoscyami. Этот гриб всюду встречается на белене. Обнаруживается он на растениях, начиная с фазы всходов, вызывает хлороз и сморщивание листьев, общее угнетение растений. Гриб в сильной степени поражает белену, снижая лекарственную ее ценность, в частности уменьшает количество алкалоидов.
К вредоносным видам принадлежит и Р. arborescens, паразитирующий на растениях семейства маковых. Он распространен во всех макосеющих районах, особенно в Восточной и Юго-Восточной Европе. Гриб поражает виды маков, относящиеся к лекарственным, декоративным и техническим растениям, вызывая сморщивание и хлороз листьев, прекращение роста плодоносцев, отмирание бутонов, уродливость цветков и коробочек. При заражении листьев гриб прободает кутикулу и эпидермис, проникает в стебель, спускается в корень, поднимается вверх и заходит в цветки, а затем в семена, где и сохраняется из года в год. P. arborescens сильно воздействует на урожай лекарственного мака, обусловливая недоразвитие коробочек, зачастую гибель растений, а также снижая количество алкалоидов у пораженных экземпляров.
P. schachtii поражает свеклу, в том числе свеклу сахарную, столовую и другие виды. В странах Европы, Северной и Южной Америки, Азии он появляется ранней весной на растениях семенников безвысадочной свеклы, где образует первые в природе очаги (обычно в марте - апреле). Возникающие на этих растениях споры разносятся ветром и воздушными течениями и попадают на поля, занятые маточной свеклой и высадками, вызывая на них массовое поражение растений. В посевах наличие гриба обнаруживается уже на самых ранних стадиях развития растений. Пораженные грибом всходы легко отличить от здоровых по значительно более бледной окраске, резко выраженной курчавости листьев. Развитие растений затем приостанавливается, в жаркое время они заметно привядают. По мере роста возникают мелкие, недоразвитые, толстые и курчавые листья с завернутыми книзу краями и с более бледной окраской пятен различной величины. Растения часто погибают.
Интенсивное развитие гриба наблюдается в период начала плодоношения растений - стрелкования (в мае - июне). В летний засушливый период активность гриба может затухать, но осенью, с изменением погодных условий, с повышением влажности, понижением температуры, развитие гриба возобновляется и на вновь образовавшихся листьях растений появляется свежее спороношение. Летнее спороношение развивается обычно на нижней стороне листовых пластинок в виде серовато-бурого с фиолетовым оттенком войлочного налета. Во влажных условиях при 25 СС зрелое спороношение образуется через 12-15 ч., но при 4 °С для того же процесса требуется более полутора суток. В сухом воздухе при 1-4 °С конидии сохраняют жизнеспособность до двух месяцев. Попадая в почву, конидии теряют способность к прорастанию через 3-6 дней под воздействием почвенных биотических и абиотических факторов, среди которых не последнюю роль играют населяющие эту среду виды микроскопических сапрофитных грибов, бактерий, животных. В течение вегетационного периода гриб образует во всех органах растений - листьях, стеблях, цветках, семенах - ооспоры. Однако прорастания последних не наблюдалось. Мицелий гриба зимует в растениях безвысадочной свеклы, в корнях маточной свеклы, в семенах.
P. schachtii вызывает преждевременное усыхание и отмирание в поле большого количества растений, приводит к значительному снижению урожая корнеплодов и их сахаристости, уменьшению урожая семян, уменьшению абсолютной массы, энергии прорастания, всхожести семян и к полному бесплодию цветоносов.
Пероноспора шпината (P. spinaceae) паразитирует на растениях семейства маревых (в частности, на шпинате), вызывая сильное поражение, а зачастую и гибель растений.
P. manshurica вызывает сильное поражение растений сои во всех зонах ее выращивания. Гриб впервые был замечен в Индии, а затем в других странах Азии (Японии и Китае), впоследствии в США и Канаде, в странах Европы (Англии, Венгрии, Польше, Румынии, Швеции, Югославии); в СССР встречается на Дальнем Востоке, Северном Кавказе, на Украине, в Московской области.
Появляется гриб весной на семядольных листьях растений сои, с которых переходит на настоящие листья, бобы, семена, вызывая поражение всего растения. Заражение других растений в поле происходит в течение всего вегетационного периода, чему способствуют повышенная влажность воздуха и умеренная температура. Жаркая и сухая погода неблагоприятна для паразита. Мицелий и ооспоры сохраняются в семенах. Ооспоры сохраняются в жизнеспособном состоянии в репродуктивных органах растений более полутора лет, а в остатках вегетативных частей в природе - до одного года.
P. brassicae поражает капусту, хотя и не причиняет этой культуре такого хозяйственного вреда, как другие виды пероноспоры: заболевают только нижние старые листья на взрослых растениях. Но при нападении на капусту в рассадный период гриб может принести культуре существенный ущерб и непосредственным воздействием на растение. Он облегчает проникновение в него факультативных паразитов, что ведет к уменьшению лежкости кочанов.
P. destructor вызывает заболевание репчатого лука (Allium), лука-батуна и других видов. Гриб очень вредоносный, широко распространенный (карта 1). Пораженные листья и цветоносы плохо развиваются, искривляются (табл. 8). Гриб появляется в период выпадения осадков и часто развивается вместе с грибом из рода стемфилий Stemphylium allii, вызывающего черную плесень лука.
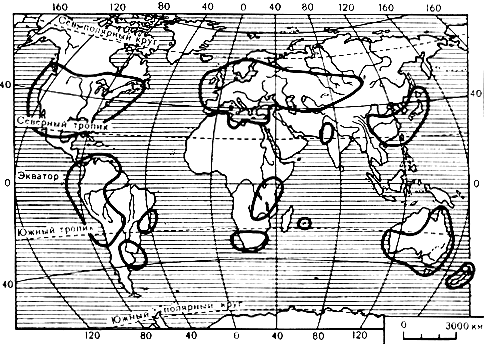
Карта 1. Ареал возбудителя ложной мучнистой росы лука
Семейство Цистоповые, или Альбуговые (Сystopaceae) (Н. С. Новотельнова)
Семейство цистоповых грибов резко обособляется среди других рассмотренных семейств порядка пероноспоровых необычным строением органов спороношения и характером паразитирования на цветковых растениях. Поражая стебли, листья, все части цветков, цистоповые вызывают сильное изменение их формы - искривление, вздутия, уродливость. Внешний вид растения настолько видоизменяется, что трудно узнать даже общеизвестную пастушью сумку, пораженную грибом Cystopus candidus.
Воздействие гриба сказывается не только на нарушении общего облика растения и отдельных его частей, но и на целостности покровов. Под влиянием паразита поверхностные ткани пораженных органов покрываются волдырями, образуя так называемые пустулы. По аналогии со ржавчиной, которая также характеризуется образованием пустул, поражение цистоповыми грибами именуют белой ржавчиной, так как споровая масса, высыпающаяся из пустул, не ржавого, а белого цвета.
Семейство цистоповых содержит один род - цистопус, или алъбуго (Cystopus), который широко распространен на земном шаре. Грибы этого рода способны легко приспосабливаться к разнообразным условиям среды благодаря своему образу жизни, своеобразному "уходу" от неблагоприятных внешних воздействий под защиту покровных тканей хозяев. Поэтому они хорошо переносят суровую зимнюю стужу и зной и сушь летнего периода, встречаются повсеместно и обнаруживаются как в северных, так и в южных странах.
Цистоповые принято рассматривать в качестве древней, прочно сложившейся группы грибов. Свидетельством того являются консерватизм и единообразие их морфологии. При этом у грибов, паразитирующих на цветковых растениях, далеко стоящих в эволюционном отношении семейств, с трудом удается уловить морфологическую неоднородность (лишь один-два нерезко выраженных признака). На примитивность цистоповых грибов указывает сохранившаяся у них обязательная стадия развития, включающая подвижные зооспоры, нуждающиеся в наличии капельно-жидкой влаги.
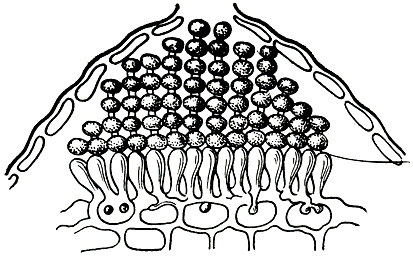
Рис. 35. Спороношение цистопуса в тканях пораженного растения
Непосредственные эволюционные связи рода Cystopus с другими низшими грибами не установлены. Возможно, что переходные формы в процессе исторического развития выпали из филогенетического ряда. Характерное для грибов этого рода спороношение состоит из слоя тесно сплоченных конидиеносцев, напоминающего гимениальный слой у высших грибов, и у других низших грибов не встречается. В то же время у водных грибов, например в семействах лептомитовых и бластокладиевых, обнаруживается "скученность" спорангиеносцев - прогрессивный признак, ставший, возможно, исходным в возникновении способности организмов к множественному спороношению вместо одиночного.
Известно около 10 видов цистоповых грибов, которые приурочены к отдельным семействам высших растений. Виды эти сложные, распадающиеся на морфологические разновидности и биологические формы. Несмотря на древность их происхождения, у них и в наши дни наблюдаются процессы дифференциации видов.
Довольно толстый неклеточный мицелий цистоповых грибов пронизывает все органы и ткани тех травянистых растений, на которых паразитирует.
Мицелий внедряется в полости клеток пораженных растений с помощью гаусторий, которые у этих грибов представляют собой пузыревидно-вздутые короткие ветви гиф, суженные в местах прохождения через клеточные оболочки. Вскоре, после того как мицелий обильно распространится внутри тканей, он переходит от вегетативного роста к размножению, бесполому и половому. Бесполые споры возникают не на поверхности органов растений, как у других пероноспоровых, а между эпидермисом и кутикулой, используя последнюю как барьер, защищающий гриб от внешних воздействий.
В процессе спороношения цистоповый гриб формирует плотные слои бесцветных толстых булавовидных клеток - спорангиеносцев, упирающихся своими вершинами в кутикулу и отрывающих ее от эпидермиса (рис. 35). На них развиваются длинные цепочки крупных, округлых, бесцветных клеток, надавливающих, в свою очередь, на кутикулу пораженного органа, вызывая трещины в ней и разрывы. В последующем целостность цепочек нарушается. Клетки отделяются друг от друга и, как тончайшая светлая пыль, высыпаются через прорванную кутикулу наружу. Эти клетки попадают с помощью ветра или брызг воды на другие растения и, осуществляя свою функцию размножения, становятся зооспорангиями. Они прорастают, образуя 4-12 зооспор, которые выходят из зооспорангиев и заражают новые растения. Однако мицелий и споры бесполого размножения не способны долго существовать и переносить суровые зимы. Сохранение гриба в природе происходит в состоянии зимних спор - ооспор, возникающих в результате полового процесса, к которому гриб переходит вскоре после осуществления бесполого размножения. На том же мицелии, проходящем в межклеточных ходах, формируются женские половые органы (шаровидные оогонии) и оплодотворяющие их мужские (булавовидные антеридии).
Ооспоры возникают по одной в каждом оогонии. Они имеют разнообразную структуру внешней оболочки - сетчатую, бородавчатую, шиповатую. Ооспора зимует и при благоприятных условиях прорастает с образованием зооспор или толстой ростковой трубки с вздутием на вершине, в котором формируется 40-60 зооспор. Но часто везикулы не развиваются, и в этих случаях ооспора прорастает и образует зачаточный мицелий. Таким образом обеспечивается ежегодное возобновление и циркуляция цистопового гриба в природе.
Самый обычный и широко распространенный вид цистоповых - Cystopus candidus (табл. 8). Он вызывает уродливость у растений свыше 240 видов крестоцветных, нападая как на дикорастущие (вайду, гулявник, желтушник, клоповник, крупку, резуху, сердечник, сирению, сумочник - табл. 8,4, сурепицу и др.), так и на культивируемые (брюкву, горчицу черную и сарептскую, капусту брюссельскую, кочанную, савойскую, цветную, кольраби, левкой, рапс, рыжик и др.). Гриб встречается на всех континентах, не только в умеренной зоне, но и в условиях юга и Крайнего Севера, в долинах и высоко в горах, с ранней весны и до поздней осени. С. candidus содержит морфологические разновидности (var. Candida и var. macrospora) и ряд специализированных форм, приуроченных к разным видам растений.
Широко распространенный гриб С. tragopogonis паразитирует на растениях семейства сложноцветных (бессмертник, бодяк, василек, девясил, пиретрум, полынь, злостный сорняк амброзия и др.), как дикорастущих, так и культурных; в Советском Союзе на юге часто встречается на козлобороднике. В 1961 г. этот гриб был неожиданно обнаружен в Краснодарском крае на подсолнечнике.
Из других видов цистопуса можно указать на С. portulacae, паразитирующий на портулаковых, С. bliti - на ширице и растениях семейства амарантовых, С. swertiae - на трипутнике.
|
ПОИСК:
|
При использовании материалов проекта активная ссылка обязательна:
http://gribochek.su/ 'Библиотека о грибах'