Изменения в составе плодовых тел грибов после отделения от мицелия
Изучение биохимических и других изменений, происходящих в грибах после сбора, имеет важное значение для рациональной организации переработки заготовленного сырья, а также для решения практических вопросов, связанных с транспортировкой и реализацией продукции в свежем виде. В литературе имеются указания, что грибы необходимо подвергать немедленной переработке или в крайнем случае хранить всего 2-3 ч (Андрест, 1968, и др.). Некоторые этот срок определяют в пределах 4-5 ч. Однако, специальных исследований по этому вопросу не проводилось. Поэтому нами на протяжении ряда лет совместно с Е. Д. Сусловой и В. А. Папилиной проводились исследования по изучению изменения качества грибов при кратковременном хранении.
Изменение органолептических показателей. В результате проведенных исследований установлена зависимость продолжительности хранения грибов от температуры окружающей среды (рис. 1). При температуре 0°С трубчатые грибы с отклонением ±15% хорошо хранятся в течение 3 сут, пластинчатые - 4-5 (опята - почти 4, а лисички - 5 сут). Во всех видах грибов за указанный период хранения не произошло внешне заметных изменений по сравнению с первоначальным состоянием. Они имели характерный внешний вид, цвет, запах и структуру. При температуре +10° С в течение первых суток хранения также не было отмечено существенных изменений в органолептических показателях. Только к концу вторых суток хранения в трубчатых грибах отмечено незначительное подсыхание краев шляпки и легкое увядание, являющееся следствием падения тур-горного давления в клетках плодового тела. Пластинчатые грибы и лисички оставались свежими.
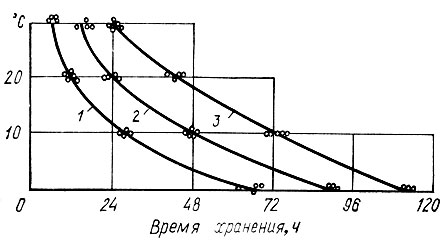
Рис. 1. Зависимость качественного состояния свежих грибов от температуры и продолжительности хранения (область влево от кривых характеризует высокое качество грибов): 1 - трубчатые грибы; 2 и 3 - пластинчатые соответственно опята и лисички
При температуре 20° С к 24 ч хранения у трубчатых грибов на поверхности шляпки и гименофора появляются коричневые пятна, а в запахе наряду с грибным чувствуется неприятный гнилостный оттенок.
Наиболее интенсивные изменения органолептических показателей происходят при температуре 30° С. Для всех видов грибов характерно появление признаков порчи в течение одних суток, причем для трубчатых грибов эти изменения наступают уже через 10-12 ч с момента отделения плодового тела от мицелия. Следует отметить, что с повышением температуры хранения время наступления признаков порчи уменьшается.
Активность ферментов и интенсивность дыхания. В плодах и овощах после уборки вследствие нарушения физиологической связи с материнским растением резко повышается активность окислительно-восстановительных ферментов. Поэтому логично предположить, что в плодовом теле грибов после отделения от мицелия должны происходить аналогичные явления, которые в свою очередь должны оказывать существенное влияние на течение биохимических процессов при кратковременном хранении и переработке.
Основным биохимическим процессом, происходящим в грибах после отделения от мицелия, является процесс дыхания, который в известной мере определяет весь ход обмена веществ в тканях. Так как на дыхание расходуются ценные питательные вещества, то общий уровень дыхания в период кратковременного хранения должен быть возможно более низким. Поэтому важно знать, как протекает процесс дыхания у отдельных грибов и как влияют на него различные факторы внешней среды.
Результаты исследования показали, что грибы после отделения от мицелия характеризуются высокой активностью дыхания, особенно опята, у которых поглощение кислорода плодовым телом составляет 325 мкл O2 на 1 г сырого вещества за 1 ч (рис. 2). Значительно меньше активность дыхания у черных груздей, белых грибов и маслят (168,9; 162,5 и 148,2), наиболее низкая - у рыжиков и лисичек.
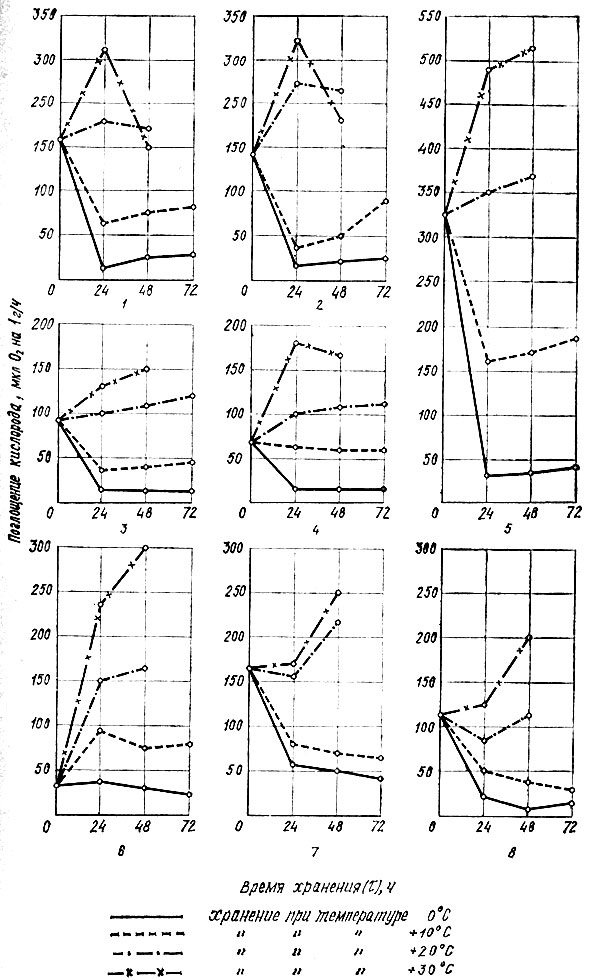
Рис. 2. Изменение интенсивности дыхания плодовых тел грибов в зависимости от температуры хранения: 1 - белых грибов; 2 - маслят; 3 - подгруздка белого; 4 - лисичек; 5 - опят; 6 - рыжиков; 7 - черных груздей; 8 - свинушек
Высокую активность дыхания опят, очевидно, можно объяснить специфическими особенностями обмена веществ данного вида грибов. В отличие от других грибов опята являются дереворазрушающими, поэтому им приходится преодолевать сопротивление защитного механизма растений, чтобы использовать содержимое клеток растения-хозяина.
Следует отметить, что в грибах интенсивность дыхания во много раз выше, чем в других растительных организмах. В частности, интенсивность дыхания овощей и плодов колеблется в среднем от 3,3 до 16,1 мкл O2 на 1 кг за 1 ч, что в десятки раз меньше, чем в свежих грибах (31,2-325 мкл O2 на 1 г за 1 ч).
После сбора интенсивность дыхания у отдельных видов грибов изменяется по-разному и зависит от температуры и длительности хранения. В маслятах и белых грибах поглощение кислорода в процессе дыхания при температуре 0°С через 24 ч составляет 18,9-21 8 мкл O2 на 1 г сырого вещества за 1 ч и на таком же уровне остается в течение 3 сут с незначительным повышением к концу хранения (для белых грибов - 2,4, для маслят - 5,6 мкл O2).
При температуре 10° С дыхание маслят и белых грибов увеличивается в 2-3 раза и быстро возрастает в период хранения. Последнее особенно выражено в маслятах, у которых после 48 ч хранения только за последующие сутки интенсивность дыхания увеличивается почти в 2 раза.
При повышенной температуре процессы дыхания в маслятах и белых грибах возрастают только в первые 24 ч хранения, после чего снижаются. Это является, очевидно, следствием развития автолитических процессов.
В подгруздке белом и лисичках, отличающихся от других грибов лучшей сохраняемостью, колебания интенсивности дыхания менее выражены и зависят от температуры и длительности хранения. Так, при 0° С у этих грибов дыхание остается ровным до конца опытного хранения, а при 10° С у лисичек наблюдается даже некоторое его снижение. У черного груздя при пониженной температуре интенсивность дыхания снижается до конца опытного хранения.
Интересно, что у пластинчатых грибов не отмечено резкого снижения интенсивности дыхания при повышенной температуре до конца хранения. Так, у рыжиков, черного груздя, свинушек и опят при температуре 30° С наблюдается повышение интенсивности дыхания в течение 48 ч. Опята, характеризующиеся самой высокой интенсивностью дыхания после сбора, отличаются и наибольшим размахом колебания данного показателя в зависимости от температуры хранения. При температуре 0° С дыхание опят к концу опытного хранения интенсивнее в 1,5-3 раза дыхания других видов грибов, при 10° С интенсивность дыхания у них находится на таком же уровне, как у подгруздка белого и лисичек при 30° С.
Дыхание является комплексным процессом, в котором участвуют многие ферментные системы. Поэтому, анализируя характер его изменения в зависимости от температуры, особенно обращая внимание на большой разрыв в интенсивности дыхания между 10 и 20° С, можно теоретически предположить, что температурный оптимум развития плодового тела грибов находится именно в этих пределах и дальнейшее уточнение замечаний закономерности и ее связь с метеорологическими условиями могут стать важной предпосылкой для объективного прогнозирования урожая дикорастущих грибов.
В период кратковременного хранения при 0°С в грибах наряду с понижением интенсивности дыхания происходит существенное уменьшение активности окислительно-восстановительных ферментов. Особенно заметно снижается активность аскорбиноксидазы (в 3-4 раза по сравнению с первоначальной). Более устойчивой оказывается монофенолмонооксигеназа (полифенолоксидаза). Увеличение срока хранения при температуре 0° С практически не оказывает влияния на изменение активности окислительных ферментов трубчатых грибов, в то время как у опят активность полифенолоксидазы за 72 ч хранения достигает почти первоначального уровня.
При температуре хранения 10° С наблюдается сначала незначительное снижение, а затем повышение активности окислительных ферментов в белых грибах и маслятах, у опят она увеличивается почти в 3 раза.
Несколько иной характер изменения активности ферментов, по-видимому, из-за видовых особенностей грибов наблюдается при повышенных температурах хранения. Так, у белых грибов при температуре 20° С в первые сутки хранения происходит снижение активности полифенолоксидазы. Затем, по мере увеличения продолжительности хранения, активность аскорбиноксидазы резко возрастает, достигая к концу опытного хранения 136,4 мкл O2 в 1 г за 1 ч. Объяснить данное явление пока еще довольно трудно. Особенно трудно объяснить изменение активности аскорбиноксидазы вследствие разнообразия ферментных систем, способных окислять аскорбиновую кислоту.
При 30° С до начала автолитических процессов у всех грибов в той или иной степени наблюдается увеличение активности окислительно-восстановительных ферментов. Особенно быстрое увеличение происходит у опят. За первые сутки хранения при 20° С активность аскорбиноксидазы возрастает в 1,6 раза, полифенолоксидазы - в 4,2 раза, при 30° С - соответственно в 3,2 и 4,5 раза. Исключение составляют свинушки, подгруздок белый и лисички. У свинушек активность полифенолоксидазы в период хранения снижается, а у подгруздка белого и лисичек при температуре 20°С резкого увеличения активности ферментов не наблюдается. Вероятно, этим можно объяснить их большую стойкость в период хранения.
Протеолитическая активность обнаружена только у опят, в виде следов у белых грибов. По мере увеличения температуры и продолжительности хранения происходит довольно значительное повышение активности протеолитических ферментов, особенно при температуре 20° С для трубчатых грибов и 30° С - для опят.
Таким образом, исследование влияния температуры на активность протеолитических, окислительно-восстановительных ферментов и интенсивность дыхания, а, следовательно, на скорость биохимических процессов показывает, что температура является решающим фактором для хранения съедобных грибов в свежем виде.
Изменения в составе основных питательных веществ. Изучение характера изменения азотистых веществ в зависимости от условий хранения имеет практическое значение и представляет теоретический интерес для выявления общих закономерностей биохимических процессов, происходящих в низших растениях.
Прежде всего необходимо отметить, что при кратковременном хранении в грибах происходит увеличение количества общего азота. Особенно большой прирост азота наблюдается при повышенной температуре хранения в трубчатых грибах. Так, при 30° С за 48 ч содержание общего азота в белых грибах увеличивается на 21, а в маслятах на 35% по отношению к содержанию на начало хранения. Интенсивность нарастания общего азота определяется в основном видовыми особенностями грибов. Характер происходящих изменений в комплексе азотистых веществ наиболее отчетливо прослеживается на примере маслят (табл. 10).
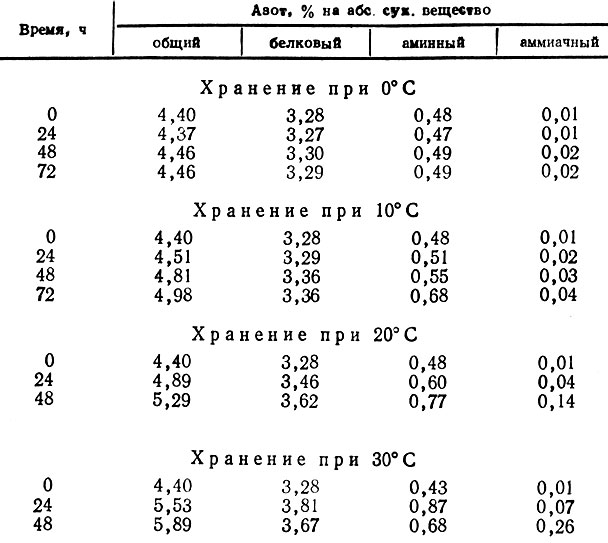
Таблица 10
При температуре хранения 0°С практически отсутствуют какие-либо изменения по всем формам азота, что согласуется с органолептическими показателями, отсутствием токсических веществ, очень низкой активностью ферментных систем и интенсивностью дыхания. Хранение при 10° С не вносит существенных изменений в содержание общего и других форм азота. При повышенной температуре скорость накопления азотистых веществ возрастает.
Значительный прирост содержания азотистых веществ в грибах после отделения их от мицелия казался трудно объяснимым. Поэтому для проверки полученных данных нами была проведена дополнительная серия опытов. Полученные данные подтвердили тенденцию увеличения всех форм азота в плодовых телах грибов в процессе кратковременного хранения при повышенной температуре. Данное явление было легче всего объяснить уменьшением количества сухих веществ, в первую очередь Сахаров, в результате усиленного дыхания. Однако, как показали расчеты, потеря Сахаров в количестве 2-3% на сухое вещество должна увеличивать процентное содержание общего азота всего на величину, в 10-15 раз меньшую, чем получалось в результате многочисленных анализов.
Из литературы известно, что некоторые грибы и микроорганизмы способны фиксировать атмосферный азот (Лилли, Барнет, 1953; Беккер, 1969; Львов, 1967; Наплекова, 1970). По мнению Любимова В. Н., процесс фиксации атмосферного азота некоторыми видами грибов сопровождается синтезом аминокислот. Поэтому есть основание предполагать, что исследованные нами плодовые тела грибов способны фиксировать азот атмосферы. Возможно, и строение плодового тела, имеющего большую поверхность соприкосновения с атмосферой, направлено на осуществление данного процесса. Высказанное предположение косвенно подтверждается отсутствием протеолитической активности на протяжении первых суток хранения.
Следует отметить, что вопрос о возможности фиксации атмосферного азота грибами интересовал ученых (Мишустин, Шильникова, 1968), но до последнего времени оставался не решенным из-за противоречивых данных. Однако последние работы чехословацких ученых (Ginterowa, 1973), работавших с мицелием съедобных грибов, подтверждают нашу точку зрения по данному вопросу. Естественно, полное доказательство фиксации атмосферного азота съедобными грибами значительно обогатит биологическую науку в раскрытии взаимосвязи между отдельными элементами леса.
Известно, что при хранении в результате преобразования аминокислот в продуктах с высоким содержанием азотистых веществ могут образовываться ядовитые вещества, подобные рыбным или трупным ядам (Ячевский, 1933, и др.). Иноуэ Идзо (Inoue, 1961) показал, что при порче съедобного гриба Armilaria matsutaka в нем полностью исчезают фенилаланин и гистидин, кроме того, обнаружены и идентифицированы фенилэтиламин и гистамин.
В наших исследованиях была поставлена задача выяснить, в каком направлении протекает процесс изменения свободных аминокислот при кратковременном хранении. В результате установлено, что в первые 24 ч хранения при +20° С общее содержание свободных аминокислот в маслятах увеличивается до 27,6%. С увеличением срока хранения начинается снижение содержания свободных аминокислот, по-видимому, в результате развития автолитических процессов. В изменениях состава и количества отдельных аминокислот трудно выявить определенную закономерность, тем не менее можно констатировать, что во всех грибах в период кратковременного хранения происходит резкое увеличение глютаминовой кислоты.
В настоящее время получила подтверждение гипотеза С. Н. Виноградского о путях фиксации атмосферного азота клубеньковыми бактериями, согласно которой первичным продуктом связывания азота является аммиак. Последний, вступая в реакцию с α-кетаглютаровой кислотой, дает глютаминовую кислоту. Глютаминовая кислота подвергаясь декарбоксилированию, переаминированию и другим превращениям, может дать другие аминокислоты. Предполагается, что такая схема связывания азота является общей для многих микроорганизмов (Кретович, 1971). Это можно рассматривать как еще одно доказательство возможности фиксации атмосферного азота шляпочными грибами.
По другим аминокислотам закономерностей изменения, характерных для всех грибов, не наблюдается. Это связано, по-видимому, с видовыми особенностями изучаемых грибов.
Потеря питательных веществ в плодовом теле грибов в период кратковременного хранения происходит в основном за счет углеводов. Это вполне объяснимо, если сопоставить полученные результаты с интенсивностью дыхания грибов после отделения плодового тела от мицелия. При дыхании в начале хранения в процесс вовлекается в основном имеющийся запас Сахаров, особенно моносахаридов. На последующих стадиях хранения, когда чувствуется недостаток моносахаридов, клетки начинают использовать сахароспирты и гликоген.
Заметное снижение активности дыхания при 0°С (на примере свинушек) характеризуется незначительным снижением общего количества Сахаров на всем протяжении трехдневного периода хранения (табл. 11).
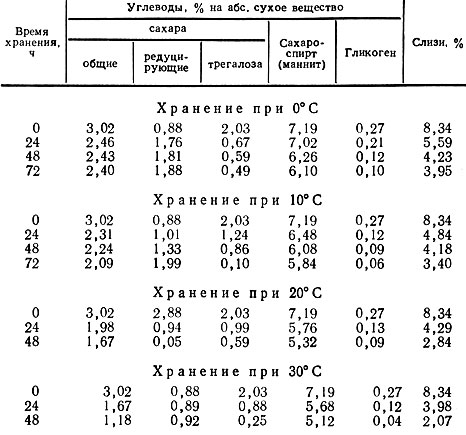
Таблица 11
Однако внутри углеводного комплекса наблюдается гидролитический распад трегалозы, гликогена (особенно на 2-е и 3-й сутки хранения) и существенное увеличение количества редуцирующих Сахаров.
С повышением температуры хранения происходит усиление процесса дыхания и резкое уменьшение общего количества сахара в основном за счет преимущественного использования трегалозы. Ее количество за 72 ч хранения при температуре 10° С уменьшается примерно в 20 раз. В период хранения при температуре 20° С по сравнению с первоначальным содержанием в результате некоторого повышения активности дыхания расход углеводов увеличивается. Эта тенденция сохраняется также и в период хранения грибов при 30° С. При указанных температурных режимах в течение 48 ч хранения практически полностью исчезает гликоген.
Интересно отметить, что в зависимости от режима хранения, например в свинушках, происходит уменьшение от 14 до 30% маннита и в 2,5-4 раза слизистых веществ. Резкое уменьшение количества слизистых веществ указывает на то, что углеводы, освобождающиеся при гидролитическом распаде слизей, активно участвуют в акте дыхания.
Таким образом, на характер потерь питательных веществ в плодовом теле грибов оказывает существенное влияние режим кратковременного хранения и Еидовые особенности грибов.
Влияние условий кратковременного хранения на структуру плодового тела. Особое значение в последнее время приобретает проблема биологических структур при совершенствовании и разработке новых методов переработки сырья растительного и животного происхождения.
Как указывалось выше, плодовое тело грибов состоит из переплетенных особым образом нежных гиф, поверхность которых непосредственно соприкасается с внешней газовой средой. Естественно, что после отделения плодового тела от мицелия в нем под действием внешних факторов должно происходить изменение механических свойств ткани, что неизбежно оказывает существенное влияние на характер испарения влаги при сушке съедобных грибов, а также на качество готового продукта.
Для характеристики изменения механических свойств структурных элементов плодового тела нами принята плотность тканей, выраженная величиной усилия (Р) в граммах, способного погрузить конический пуансон с углом при вершине 60° на глубину 1 см в течение 3 мин.
На рис. 3 показано влияние условий и продолжительности хранения грибов на изменение плотности тканей плодового тела.
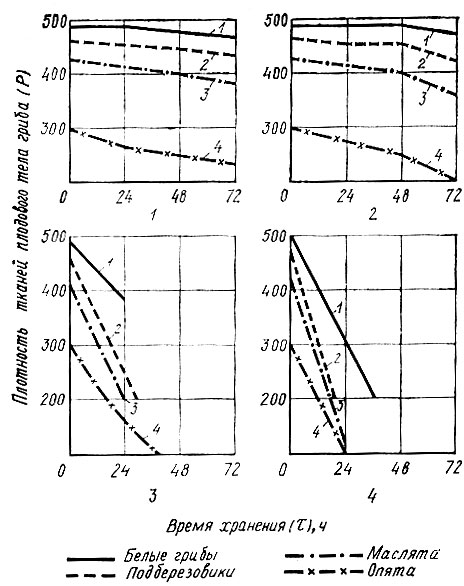
Рис. 3. Влияние условий и продолжительности хранения на изменение плотности тканей плодового тела: 1 - хранение при 0° С; 2 - хранение при 10° С; 3 - хранение при 20° С; 4 - хранение при 30° С
Из представленных графиков следует, что температура хранения оказывает существенное влияние на характер изменения плотности и структуры тканей. Так, при 0°С свежие грибы на протяжении 3 сут хранения сохраняют механические свойства структурных элементов плодового тела примерно на одинаковом уровне, за исключением опят, у которых на 3-4 сутки плотность тканей снижается примерно на 15%. Несколько существеннее изменяется плотность тканей при хранении грибов при 10° С. При повышенной температуре хранения в результате усиленных биохимических процессов наблюдается стремительное снижение плотности тканей, что согласуется с данными по увеличению активности ферментов, интенсивности дыхания, изменениям в комплексе азотистых веществ и углеводов.
Развитие личинок насекомых в зависимости от условий хранения. Плодовое тело грибов в период своего развития поражается многими видами вредителей. По данным Островерховой-Плотниковой Г. И. (1964), в условиях Западной Сибири белые грибы поражаются 22 видами насекомых, маслята поздние - 19 видами и т. д. В неблагоприятные годы степень поражения может достигать 89,4%. По данным Б. П. Василькова (1968), а также И. И. Журавлева, средняя пораженность составляет до 50%. В. И. Шубин (1976) определяет ее в пределах 30-70% в зависимости от погодных условий.
Наличие личинок насекомых снижает качество грибного сырья и готовой продукции. Однако добиться полного исключения случаев наличия червоточин или самих личинок практически нельзя. A. binder, К. Löweneck (1964) показали, что даже при самой тщательной сортировке грибного сырья перед консервированием пораженность личинками кусочков грибов колеблется от 9 до 23%. Анализируя грибы теневой сушки, они также часто обнаруживали сильное поражение продукта. Нами изучалась интенсивность развития личинок в плодовых телах грибов после отделения их от мицелия в зависимости от степени поражения, продолжительности и температуры хранения.
Исследования показали, что количество личинок в маслятах при одинаковом поражении ножки (2-4 червоточины) колеблется в широких пределах. Так, грибов со слабой пораженностью (до 100 личинок на 100 г грибов) было 32%, со средней (101-1000) - 52% и сильной (1001-2000) - 16%. Отсюда следует, что степень пораженности ножки не может быть критерием для определения степени пораженности всего плодового тела.
При благоприятных для развития личинок насекомых условиях хранения происходит стремительное снижение качества грибов.
Наблюдения за хранением грибов при 20° С показали, что грибы со слабой пораженностью могут подвергаться кратковременному хранению в пределах 20 ч, а грибы с более высокой степенью пораженности необходимо после тщательной сортировки подвергать немедленной переработке.
Решающее влияние на интенсивность развития личинок насекомых оказывают температура и продолжительность хранения (табл. 12).
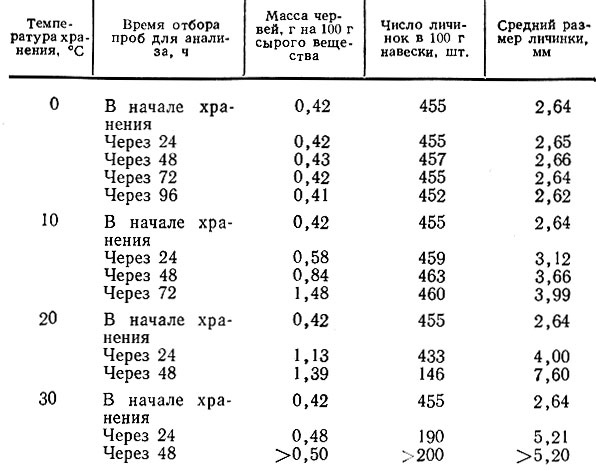
Таблица 12
При 0°С масса и средний размер личинок насекомых в течение 4 сут хранения практически не изменяется. К концу хранения личинки становились малоподвижными, тощими, некоторая часть их погибала.
При 10° С развитие личинок происходит довольно быстро. Через 72 ч масса их увеличивается в 3,5 раза, отмечено увеличение размера и числа их за счет появления новых личинок. Средний размер личинок (2,62-7,6 мм).
Как уже отмечалось выше, исключительно быстро происходит развитие насекомых при температуре 20° С, а еще быстрее при 30° С, что практически приводит к полному уничтожению грибов уже через сутки.
Полученные нами данные показывают, что при повышенной температуре хранения с каждым часом происходит увеличение количества личинок и уничтожение плодовой ткани грибов. Поэтому на перерабатывающих предприятиях грибы до переработки следует хранить при температуре, близкой к 0°С, но не выше 10° С.
Таким образом, после отделения плодовых тел от мицелия основным фактором, влияющим на интенсивность изменения качества грибного сырья, является температура. С повышением температуры происходит увеличение активности ферментных систем и интенсивности дыхания, что вызывает значительные изменения в комплексе азотистых веществ, уменьшение количества углеводов и снижение органолептических показателей.
Принимая во внимание закономерности изменения биохимического характера органолептических свойств и структуры плодового тела, можно сделать вывод, что зависимость качества от температуры и продолжительности хранения (см. рис. 1) может служить для определения сроков хранения грибного сырья до переработки в конкретно сложившейся обстановке на предприятии.
|
ПОИСК:
|
При использовании материалов проекта активная ссылка обязательна:
http://gribochek.su/ 'Библиотека о грибах'