5. Ядро
Ядерная оболочка грибов (Piriculariae oryzae, Rhizopus nigricans) двухслойная, толщиной 200 А, снабженная порами диаметром 600-800 А. У метаболирующего ядра вегетативных гиф ядерная оболочка складчатая, что, вероятно, связано с увеличением его поверхности в процессе адаптации к активному обмену с цитоплазмой (Хакимова, 1971; Necas et al., 1963). Ядерные поры грибов имеют гранулярную или фибриллярную структуру (Фрей-Висслинг, 1976). В полости ядра, например, Penicillium chrysogenum встречаются многочисленные гранулы и нитчатые тельца, состав и назначение которых не ясны. В ядре локализуются ферменты, участвующие в синтезе нуклеиновых кислот, ядерных белков, нуклеопротеидов и нуклеотидов. Имеются сведения о локализации в ядре центра синтеза дегидрогеназ пиридинового и флавинового типов.
Основной морфологической особенностью ядер грибов является разнообразие способов их деления и образования ядерного веретена, а также подобно инфузориям явление "ядерного дуализма".
Центриоли у большинства грибов, кроме некоторых оомицетов и зигомицетов, отсутствуют. Вместо них к оболочкам ядер прилегают гранулярные или дисковидные электронно-плотные тельца с диаметром 0,5-2 мкм, толщиной 70 мкм, называемые полярными тельцами веретена (ПТВ). Расположение их и связывающих их микротрубочек веретена в поздней анафазе деления ядра дискомицета Peziza badla показано в атласе (Камалетдинова, Васильев, 1982/рис. 26). Как ПТВ, так и микротрубочки предсуществуют в клетках сумчатых и базидиальных грибов, обнаруживаются в них в промежутках между митозами. ПТВ составлены из плотноупакованных гранул или трубочек и, видимо, самовоспроизводятся, так как содержат белок, РНК и, возможно, также ДНК (Камалетдинова, Васильев, 1982). Микротрубочки составлены из специфического белка - тубулина.
По типу образования ядерного веретена грибы можно разделить на две основные группы (рис. 7.5): 1) группа, включающая миксомицеты, оомицеты и зигомицеты, с веретеном, образующимся из ядрышка, и с формированием колец или колпачков хроматина на полюсах деления в анафазе, и 2) группа хитридиевых, базидиальных, сумчатых и несовершенных грибов с веретеном, образующимся из предсуществующих микротрубочек (Burnett, 1968), с ПТВ и исчезающим во время деления ядрышком.
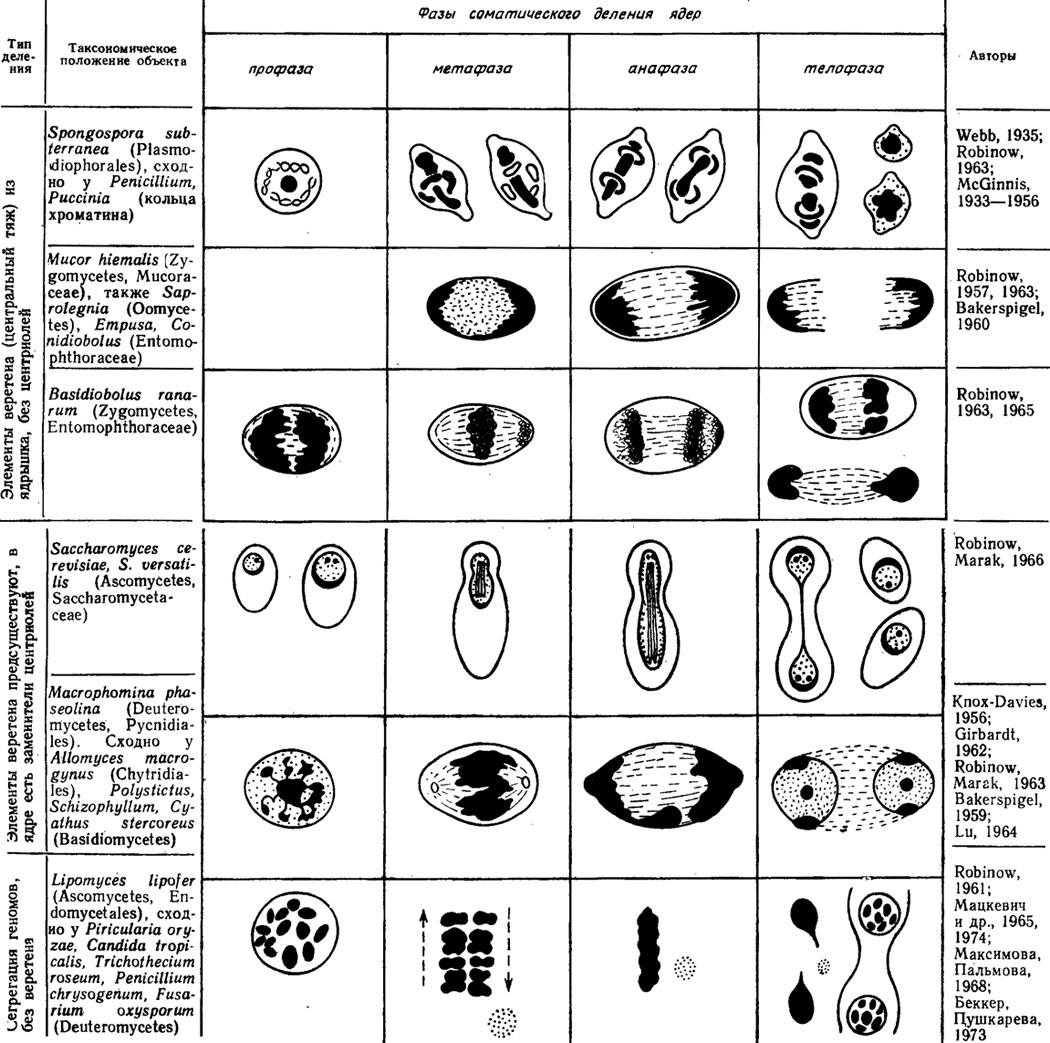
Рис. 7.5. Сводная схема типов соматических делений ядра у грибов с помощью аппарата веретена различного происхождения, способом сегрегации хромосом и скольжения геномов
Для деления ядер в вегетативных гифах характерно слипание хромосом и распределение их в метафазе по всей длине веретена, как это можно видеть на примере грибов ржавчины (рис. 7.6/Курсанов, 1915). Поэтому подсчитать их число в вегетативных образованиях практически невозможно и удается с достаточной точностью только при их четком разобщении в процессе редукционного деления (Olive, 1965). Для вегетативных структур грибов характерно параллельное существование ряда разнообразных способов деления. При соматических делениях Piricularia oryzae (Хакимова, 1971) их можно насчитать не менее пяти: классический митоз, деление путем скольжения хроматина вдоль оси, отмеченное также у Lypomyces (Robinow, 1965), путем сегрегации геномов, атипичное с образованием анеуплоидов с неравным числом хромосом и деление без расхождения геномов, ведущее к образованию полиплоидных ядер (рис. 7.7). Сходная картина разнообразных способов деления ядер была обнаружена в мицелии и конидиеносцах хищного гриба Arthrobotrys (Теплякова, 1977).
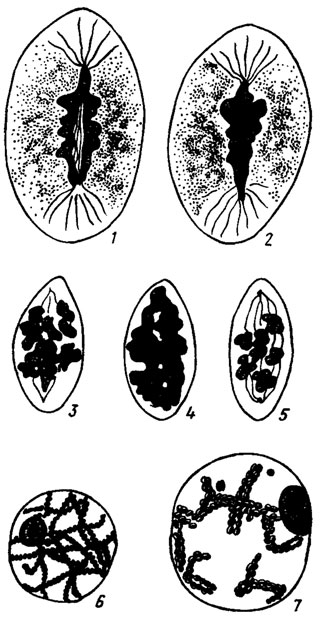
Рис. 7.6. Слияние хромосом и распределение их по длине веретена в метафазе в вегетативных гифах грибов ржавчины (1-5). Формирование бивалентов хромосом в профазе деления (6-7) (Курсантов, 1915)

Рис. 7.7. Различные типы деления ядра у Piricularia oryzae: a - деление по типу классического митоза; б - деление по типу сегрегации геномов с образованием сборных хромосом. Стрелками обозначены полярные тельца веретена (ПТВ); в - атипичный митоз; г - диплоидные и полиплоидные ядра (Хакимова, 1971)
У ряда грибов обнаружено явление ядерного дуализма. У Penicillium chrysogenum наблюдается на ранних этапах развития, а у Fusarium oxysporum и позднее два типа ядер - крупные неделящиеся, с хроматином по периферии ядра и мелкие, целиком заполненные хроматином, часто находящиеся в стадии деления (Беккер, 1956, 1975). Такая картина обнаруживается при окраске исследуемого объекта по методу Фельгена. Прижизненная окраска акридин-оранж показала, что крупные ядра - желтые, тогда как мелкие - зеленые (Беккер, Чадова, 1973), что говорит об обогащенности мелких ядер ДНК, тогда как в крупных имеется и большая доля РНК. Эта особенность ядер грибов чрезвычайно сходна с характерной для микро- и макронуклеусов инфузорий. Существование в онтогенезе грибов ядерного дуализма имеет огромное значение для их адаптивной способности и изменчивости. Формирование у грибов крупных, видимо, полиплоидных ядер, гомологов макронуклеусов, возможно, начинается, как и у инфузорий, с эндополиплоидии, которая была установлена у дрожжей и гифообразующих грибов (Мацкевич, Головня, 1965; Мацкевич, 1981). Назначение мелких генеративных ядер грибов аналогично микронуклеусам у инфузорий состоит, по-видимому, в сохранении всей генетической информации в целом.
Из других структурных элементов грибных клеток можно упомянуть о ломасомах, часто расположенных в углах соприкосновения септы с боковой клеточной стенкой, о диктиосомах, связанных с жгутиками зооспор (Громов, Мамкаева, 1975), и о лизосомах, представляющих собой часть вакуолярного аппарата клетки. Встречаются иногда в заметных количествах пигментированные элементы, осмофильные и не осмофильные липидные включения, отграниченные от цитоплазмы однослойной оболочкой (Камалетдинова, Васильев, 1982).
|
ПОИСК:
|
При использовании материалов проекта активная ссылка обязательна:
http://gribochek.su/ 'Библиотека о грибах'