2. Пути усвоения неорганического азота грибами
Общая схема усвоения нитрата грибами состоит из четырех этапов (рис. 4.1). В этой реакции участвуют промежуточные этапы переноса электронов, в которых обязательными компонентами являются молибден и флавопротеид (ФАД). Потеря нитратом двух электронов приводит к образованию нитрита, имеющего на два заряда меньше предыдущего.
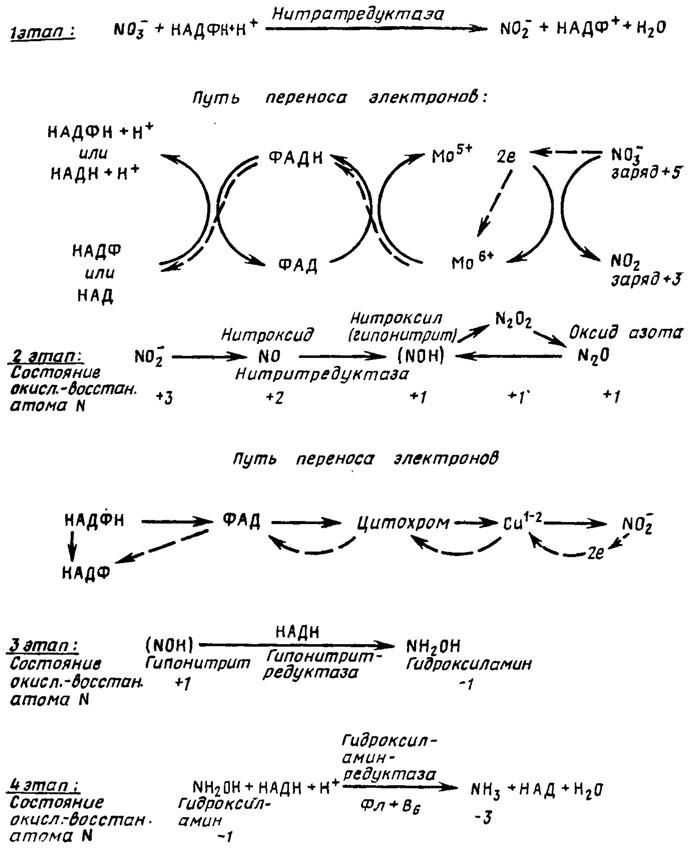
Рис. 4.1. Схема процесса усвоения нитратов грибами
Второй этап, осуществляемый ферментом нитритредуктазой, состоит в переходе с потерей молекулой еще двух электронов из нитрита в нитроксид (NО) и затем в иитроксил (NOH) или гипонитрит. На этой стадии возможен также переход без потери валентности из нитроксила (NOH) в N2O2 и далее в N2O и обратно. Наличие этого промежуточного этапа подтверждается существованием у Neurospora энзима, восстанавливающего нитрит до оксида азота (1) (Walker, Nicholas, 1961), причем у грибов показана способность усваивать это соединение (Nicholas, 1965). В процессах второго этапа восстановления неорганического азота участвуют как кофакторы НАДФ, ФАД, цитохром и ион меди.
Третий этап восстановления, протекающий с участием энзима гипонитритредуктазы, состоит в восстановлении гипонитрита до гидроксиламина. Гипонитритредуктаза была обнаружена у Neurospora, причем выяснилось, что кофактором ее является восстановленный НАД.
Четвертый этап состоит в восстановлении гидроксиламина до аммиака, протекающем с участием НАДН при действии фермента гидроксиламинредуктазы. Реакция восстановления гидроксиламина в аммиак была установлена у Neurospora. Энзим оказался так же, как и в двух первых этапах восстановления азота флавопротеином, нуждающимся в пиродоксине и в металлах, которые подавляются хелирующими агентами. Для синтеза гидроксиламинредуктазы оказались необходимыми марганец и магний, нужные также и для синтеза нитрит- и гипонитритредуктаз (табл. 4.2).
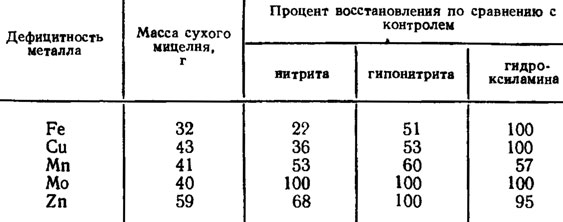
Таблица 4.2. Действие дефицита металлов на активность нитрит-, гипонитрит- и гидроксиламинредуктаз Neurospora crassa (Medina, Nicholas, 1957)
Для функции нитрит- и гипонитритредуктаз оказались необходимыми железо и медь, а молибден только для нитратредуктазы (Medina, Nicholas, 1957). Из других металлов для синтеза нитратредуктазы необходим также цинк.
Весь процесс восстановления нитрата в целом нуждается в следующих кофакторах: НАДФ (или НАД), ФАД, сульфгидрильных группах (табл. 4.3), в фосфате (сходный эффект дают также арсенит и пирофосфат), а на этапе синтеза гидроксиламинредуктазы еще в пиридоксальфосфате. В последнем этапе возможно образование комплекса (промежуточного) с гидроксиламином, в начале в форме оксима, а потом амина.

Таблица 4.3. Общая схема потребности в компонентах редуктазиой системы при восстановлении нитрата и других соединений азота у грибов (Nicholas, 1965)
Пятый этап - утилизация аммиака для биосинтеза аминокислот - протекает при участии глютаминдегидрогеназы и состоит в аминировании α-кетоглютаровой кислоты с параллельным восстановлением ее кетогруппы (R-СО-) в группу R-CHNH2. Такой энзим был найден у Neurospora crassa (Nicholas, 1965), причем было выяснено, что для его биосинтеза требуется цинк. У грибов глютаминдегидрогеназа - ключевой энзим для продукции органических соединений азота, и мутанты, не имеющие этого энзима (например, мутант Neurospora N 2003), не растут на средах с ионом аммония в качестве источника азота, а только на содержащих аминокислоты. Общая схема потребностей энзимов системы нитратредуктазы в различных кофакторах приведена в табл. 4.3.
Возросший в последнее время интерес к механизму азотфиксации прокариотными организмами привел к широкому ряду работ по общему изучению молибденсодержащих ферментов, в том числе и нитратредуктазы - первого комплекса энзимов в пути восстановления неорганического азота до аммиака, оказавшегося у прокариотов тесно связанным с фиксирующим азот воздуха ферментом нитрогеназой. При этом были затронуты и грибные организмы, в частности дрожжи Rhodotorula glutinis и дефектный по молибденовому компоненту мутант nit-1 Neurospora crassa (Ketchum et al., 1970; Ketchum, Sevilla, 1973; Pan, Nason, 1978).
Ассимиляторная нитратредуктаза эукариотов, осуществляющая двухэлектронное восстановление нитратов в нитриты, оказалась двухкомпонентным комплексом из диафоразы, содержащей как кофакторы у грибов НАДФН (Львов и др., 1980), и терминальной молибденсодержащей нитратредуктазы (табл. 4.4). Промежуточным переносчиком между ними служит цитохром b557, а вся схема переноса восстановленных эквивалентов на нитрат выглядит следующим образом:
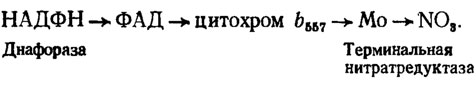
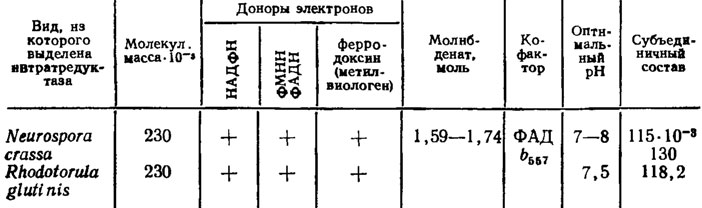
Таблица 4.4. Характеристика ассимиляторных нитратредуктаз грибов (Львов и др., 1980)
У прокариотов эта цепь ограничена двумя последними компонентами с донором электронов в форме ферродоксина или его химического аналога метилвиологена. Кофакторы молибденсодержащих ферментов оказались общими для целого ряда энзимов, таких как нитратредуктаза, ксантиноксидаза, альдегидоксидаза, сульфитоксидаза и ксантиндегидрогеназа (Маккена и др., 1974; Львов и др., 1980). У Neurospora crassa этот кофактор оказался низкомолекулярным (Ketchum, Sevilla, 1973), что подтвердилось при самосборке ее нитратредуктазы (Pan, Nason, 1978), тогда как кофактор, входящий в состав нитрогеназы прокариотов, содержит не менее восьми атомов железа и шести атомов серы на один атом молибдена (Shah, Brill, 1977). Однако молибденовый компонент их нитратредуктазы в 2-4 раза ниже по молекулярной массе, чем у эукариот, и содержит не два, а только один атом молибдена (см. табл. 4.4). Исследования по нитратредуктазам и нитрогеназе еще не закончены и продолжают пополняться новыми сведениями.
Кроме нитратов грибы способны восстанавливать также и органические нитросоединения, например м-нитрозонитробензол в м-нитроанилин. Такая реакция описана у Neurospora (Nicholas, 1965), а восстановление n-нитробензола у Aspergillus niger.
У грибов установлено так называемое азотное дыхание, т. е. окисление соединений азота, возможной целью которого является получение энергии, аналогичное подобному процессу у бактерий нитрификаторов (Nicholas, 1965). Материалом для этого окисления служит аммиак, выделяющийся в результате дезаминирования аминокислот, и обнаруживают эту способность чаще всего у почвенных гифомицетов, обитающих в условиях южных пустынных почв, бедных источниками углерода (Aspergillus flavus, A. wentiif Penicillium atrowenetum). Процесс окисления, видимо, идет у них в направлении, обратном процессу восстановления нитратов, так как среди промежуточных продуктов его обнаруживаются гидроксиламин и нитрит, а заканчивается процесс образованием нитрата. Впрочем, энергетическое значение этого процесса в настоящее время подвергается сомнению (Мирчинк, 1976).
|
ПОИСК:
|
При использовании материалов проекта активная ссылка обязательна:
http://gribochek.su/ 'Библиотека о грибах'