Микофильные несовершенные грибы (И. И. Сидорова)
Микофильные грибы, обитающие на других грибах,- обширная экологическая группа, включающая не менее 1500 видов. В нее входят представители всех классов грибов, однако наиболее широко микофилы представлены в таких группах, как некоторые хитридиомицеты, семейства пиптоцефаловых и димаргарисовых из порядка мукоровых (класс зигомицеты), порядок гипокрейных из класса аскомицетов и другие. Большое их число относится и к классу дейтеромицетов.
Микофильные дейтеромицеты широко распространены в разных климатических зонах земного шара. Многие виды из этой группы - космополиты, например хорошо известный паразит на ржавчинных грибах дарлюка нитчатая (Darluca filum). Другие обитают в умеренной зоне обоих полушарий. Особенно богато и разнообразно представлена эта экологическая группа грибов в тропиках, в условиях теплого и влажного климата.
Дейтеромицеты-микофилы встречаются на грибах из различных таксономических и экологических групп. Их можно легко обнаружить на крупных плодовых телах базидиомицетов и аскомицетов, на склероциях и стромах различных грибов, на многих грибах - паразитах растений (ржавчинных, мучнисторосяных, головневых и др.). Многие грибы, обитающие в почве, например виды из рода триходерма, ризоктония картофельная (Bhizoctonia solani), некоторые пенициллы, способны паразитировать на других почвенных грибах. В этом отношении они немного напоминают хищные грибы. Развиваясь сапрофитно в почве или на растительных остатках, микофилы получают дополнительное питание из мицелия других грибов.
По характеру взаимоотношений с грибами-хозяевами группа микофильных дейтеромицетов очень разнородна. В нее входят как паразиты, так и сапрофиты, развивающиеся преимущественно на отмерших плодовых телах грибов или на некротизированной ткани растений вокруг спороношений фитопатогенных грибов. Представители последних довольно часто встречаются в природе. На старых, высохших шляпочных грибах часто можно заметить оливковый или черновато-зеленый бархатистый налет, образованный конидиеносцами и конидиями различных грибов из рода кладоспорий. Некоторые пенициллы, например пеницилл Тома (Penicillium thomii), также часто встречаются на плодовых телах гименомицетов, образуя на них зеленовато-голубой порошистый налет. На старых эцидиях и уредопустулах многих ржавчинных грибов часто встречается темный налет мицелия и спороношений сапрофитных видов из родов альтернария, кладоспорий и др. Все эти микофилы - типичные сапрофиты, встречающиеся на разнообразных растительных субстратах или в почве и способные также заселять уже отмершие части грибов.
Микофильные грибы, паразитирующие на других грибах, называют микопаразитами. Для микопаразитов же, обитающих на фитопатогенных грибах, часто используют название сверхпаразиты или гиперпаразиты. Микопаразитизм у дейтеромицетов известен давно. А. де Бари еще в 1870 г. описал пикнидиальный гриб цицинноболюс Цезати (Cicinnobolus cesatii), паразитирующий на мучнисторосяных грибах. Однако интенсивное изучение этой экологической группы грибов было начато только в 30-х годах нашего века. Повышенный интерес к микопаразитам, особенно в последние годы, объясняется несколькими причинами. Представители этой группы - естественные враги многих фитопа-тогенных грибов, например ржавчинных и мучнисторосяных. Разработка биологических методов борьбы с болезнями растений при помощи гиперпаразитов требует их широких флористических исследований, выяснения их роли в биоценозах, в регуляции численности популяции фитопатогенных грибов. Изучение многих микопаразитов показало, что система микопаразит-хозяин - очень удобная модель для исследования основных принципов паразитизма у грибов. Они быстро растут в культуре, их легко выращивать в строго контролируемых условиях. Наконец, некоторые микопаразиты образуют антибиотики, подавляющие развитие грибов, а также ферменты, разрушающие их клеточные стенки (хитиназа, глюканазы). Один из антибиотиков микопаразитов - трихотецин - уже производит промышленность, и его применяют в растениеводстве и животноводстве для борьбы с болезнями, вызываемыми грибами. Ферменты микопаразитов в будущем смогут найти применение в промышленности для разрушения клеточных стенок грибов и повышения усвояемости грибов.
Как и грибы - паразиты высших растений, микопаразиты различаются по характеру питания. Одни из них могут питаться только содержимым живых клеток хозяина, и поэтому их называют биотрофными микопаразитами. Другие сначала убивают клетки хозяина, образуя антибиотики или ферменты, а затем питаются содержимым отмерших клеток. Эту группу микопаразитов называют некротрофными микопаразитами. Граница между этими двумя группами микопаразитов не всегда четка. Известны микопаразиты (цицинноболюс Цезати, ризоктония картофельная и др.), гифы которых внедряются в живые клетки хозяина и в течение некоторого времени существуют биотрофно, однако позднее убивают эти клетки и живут как некротрофы.
Обычно биотрофные микопаразиты довольно узко специализированы, а нектотрофы имеют широкий круг хозяев. Однако из этого правила есть исключения. Так, биотрофный микопаразит стефанома буроспоровая (Stephanoma phaeospora), обнаруженная на грибе из рода фузариум, может паразитировать на грибах из классов аскомицетов, базидиомицетов и дейтеромицетов.
Биотрофные микопаразиты преобладают среди микофильных хитридиомицетов и зигомицетов. Но лишь у немногих микофильных дейтеромицетов обнаружен биотрофный, хорошо сбалансированный характер паразитизма. Одни из них довольно широко распространены в природе, как гонатобэтрис простой (Gonatobotrys simplex), паразитирующий на альтернариях, другие встречаются редко. Например, гонатоботриум бурый (Gonatobotryum fuscum) обнаружен только в Англии и в штате Западная Виргиния в США на видах рода цератоцистис (Ceratocystis), а калъкариспориум паразитический (Calcarisporium parasiticum) - только в США на дотиорелле дубовой (Dothiorella quercina). Немногочисленность известных сейчас биотрофных дейтеромицетов объясняется прежде всего тем, что их довольно трудно обнаружить в природе и получить в культуре, где они растут только в присутствии хозяина или на средах специального состава.
У биотрофных дейтеромицетов обнаружен своеобразный характер взаимодействия гиф паразита и хозяина. В отличие от биотрофных хитридиомицетов и зигомицетов, они никогда не внедряются в клетки хозяина; их контакты с гифами хозяина всегда имеют экстрацеллюлярный (внеклеточный) характер. Это так называемые контактные микопаразиты. Они образуют абсорбтивные (питающие) гифы различного строения, устанавливающие контакт с клетками хозяина, не внедряясь в них. Механизм питания при таком контакте неизвестен. Предполагают, что в зоне контакта гиф паразита и хозяина повышается проницаемость цитоплазматической мембраны клеток последних и происходит поглощение их метаболитов клетками паразита. Строение абсорбтивных гиф различается у разных видов контактных микопаразитов. Так, у калькариспориума паразитического образуются специальные контактные, или буферные, клетки, гонатоботрис простой обвивает своими гифами гифы хозяина, образуя контактные веточки, а стефанома буроспоровая образует характерные группы контактных клеток, так плотно охватывающие гифы хозяина, что их трудно отделить даже препаровальной иглой.
В природе биотрофные паразиты не могут развиваться в отсутствие своего хозяина. Поэтому у некоторых из них в процессе эволюции выработались специальные приспособления, облегчающие контакт с его гифами. Споры калькариспориума паразитического и гонатоботриса простого не прорастают в воде и на обычных питательных средах, а только в присутствии хозяина, экстракта из мицелия грибов и некоторых растительных продуктов. У первого прорастающие споры образуют лишь очень короткие ростковые трубки, однако гифы хозяина обнаруживают положительный хемотропизм и растут по направлению к ним. Такой тропизм у гиф хозяина наблюдается при микопаразитизме очень редко. Чаще встречается другой тип тропизма - гифы паразита или ростковые трубки его спор растут по направлению к гифам или спорам хозяина. Он обнаружен у некоторых некротрофных дейтеромицетов, например дар люки нитчатой, иубиотрофных зигомицетов.
Биотрофные дейтеромицеты контактного типа не растут на обычных питательных средах в отсутствие хозяина. Для их развития необходимы витамины и ростовой фактор микотрофеин, содержащийся в мицелии многих грибов. Если к синтетической питательной среде с глюкозой добавить водный экстракт из мицелия грибов, содержащий микотрофеин, на такой среде хорошо растут калькариспориум паразитический, гонатоботрис простой и гонатородиелла Хайли (Gonatorohodiella highlii).
Как и сбалансированные паразиты высших растений, биотрофные микопаразиты не вызывают быстрой гибели хозяина, а только замедление его роста.
Большинство дейтеромицетов-микопаразитов относится к группе некротрофов или паразитов смешанного типа. У многих из них характер паразитизма пока не установлен. Некротрофные микопаразиты и микопаразиты смешанного типа, вызывающие быстрое разрушение клеток хозяина, не образуют специализированных абсорбтивных структур типа гаусторий или контактных клеток. Контакт с клеткой хозяина осуществляется у них как путем образования неспециализированных внутриклеточных гиф, так и экстрацеллюлярно, причем гифы паразита плотно обвивают гифы хозяина. Нередко у одного и того же микопаразита наблюдаются оба типа контакта с хозяином. Дарлюка нитчатая - гиперпаразит на многочисленных ржавчинных грибах - обычно образует экстрацеллюлярные гифы, но в некоторых случаях ее гифы внедряются в уредоспоры хозяев. Биохимические основы взаимодействия гиф паразита и хозяина пока не выяснены. Вероятно, внедрению гиф паразита в клетки хозяина предшествует лизис клеточных стенок последних. Многие некротрофные паразиты вызывают быстрый лизис клеточных стенок гиф хозяина. У некоторых из них обнаружены ферменты, гидролизующие полимеры клеточных стенок грибов - хитин и различные глюканы. В культурах триходермы зеленой (Trichoderma viride), например, образуются хитиназа и две глюканазы, а у кониотирия минитанс (Coniothyrium minitans), паразитирующего на склероциях грибов,- хи-тиназа и глюканазы, гидролизующие полисахарид склероциев - склеротан.
Большую роль в жизни многих некротрофных микопаразитов играют образуемые ими антибиотики. Такие грибы, как трихотециум розовый (Trichothecium roseum), глиокладиум розовый (Gliocladium roseum), вертицилл гемилейи (Verticillium hemileiae), некоторые виды из рода триходерма, образуют антибиотики, убивающие мицелий других грибов, и используют его в качестве питательного субстрата.
Некротрофные паразиты вызывают быструю гибель пораженных структур хозяина, сильно замедляют или полностью подавляют его развитие. Развиваясь на спорообразующих структурах грибов, они часто ограничивают их спороношение и снижают жизнеспособность или вызывают гибель образующихся спор. Представители этой группы обычно хорошо растут в лаборатории на питательных средах. Поэтому некротрофные микопаразиты и микопаразиты смешанного типа представляют большой интерес с точки зрения разработки биологических методов борьбы с грибами - паразитами растений. Во всех работах по биологическим средствам защиты растений с помощью гиперпаразитов фитопатогенных грибов используются микопаразиты из этой группы - триходерма зеленая, дарлюка нитчатая, цицинноболюс Цезати и многие другие.
В нашей стране исследования по биологическим методам борьбы с фитопатогенными грибами с использованием гиперпаразитов были начаты еще в 30-х годах (Н. С. Федоринчик, M. Е. Владимирская) и широко проводятся в настоящее время во многих научно-исследовательских учреждениях. В результате их установлена перспективность использования некоторых гиперпаразитов в защите растений от грибных болезней, например препарата триходермы зеленой - триходермина, паразита мучнисторосяных грибов - цицинноболюса Цезати и некоторых других. Второе перспективное направление - получение антибиотиков, образуемых микопаразитами. Один из них - трихотецин - уже нашел применение в практике.
Наиболее распространенный и хорошо известный гиперпаразит ржавчинных грибов - дарлюка нитчатая - относится к сферопсидным грибам. У него известна и сумчатая стадия - эударлюка осоковая (Eudarluca caricis), относящаяся к локулоаскомицетам. Этот гриб - космополит, встречающийся на всех континентах на различных ржавчинных грибах. Он поражает преимущественно уредостадию ржавчинных грибов, но встречается также на эцидиях и телейтопустулах. Круг хозяев у этого вида очень широк и включает ржавчинники из семейства пукциниевых и реже мелампсоровых. Однако природные популяции гиперпаразита состоят из множества штаммов, различающихся по вирулентности по отношению к различным видам ржавчинных грибов (П. Д. Кинер).
Развиваясь на пустулах ржавчинных грибов, дарлюка нитчатая образует белый, обычно малозаметный мицелий, а на нем маленькие шаровидные пикниды, диаметром 50-200 мкм, под лупой выглядящие как маленькие черные шарики (рис. 268). В одной пустуле их образуется от 1-3 до нескольких десятков. В каждой пикниде образуется около 6000-8000 спор, бесцветных, веретеновидных, с одной перегородкой и нитчатыми придатками на концах. Споры склеены слизью и выходят из пикниды в виде длинной беловатой или сероватой нити. Попадая на растения, споры прорастают в каплях воды. Если растение не заражено ржавчиной, проросшие споры погибают. Если они прорастают в пустуле ржавчинного гриба или вблизи его уредоспор, развивается мицелий паразита, окружающий гифы и споры хозяина и питающийся их содержимым. Иногда он проникает и внутрь клеток хозяина. При прорастании спор гиперпаразита в смеси с уредоспорами ржавчинных грибов у их ростковых трубок наблюдается тропическая реакция - они растут по направлению к прорастающим уредоспорам хозяина. Кроме того, прорастающие уредоспоры стимулируют прорастание конидий гиперпаразита.
Интенсивное развитие дарлюки нитчатой на ржавчинных грибах происходит при высокой относительной влажности воздуха и температуре в границах 12-26 °С. Поэтому этот вид может развиваться почти во всех районах земного шара, где встречается ржавчина, от тропиков до северных границ ее ареала. При высокой влажности воздуха гриб образует обильный воздушный мицелий, а при низкой влажности его мицелий незаметен, он часто уходит внутрь тканей растения вместе с мицелием хозяина.
Процесс формирования пикнид после заражения пустул ржавчинного гриба занимает 5-7 дней, таким образом за вегетационный период развивается несколько поколений гиперпаразита.
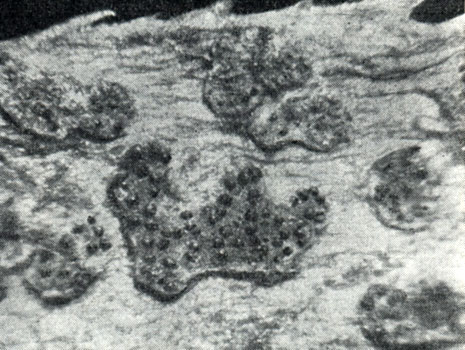
Рис. 268. Пикниды гриба дарлюка нитчатая (Darluca filum) в уредопустуле ржавчины
Зимуют пикниды гриба или его сумчатая стадия на растительных остатках. Жизнеспособность перезимовавших в пикнидах спор гриба различна в разных районах. Так, выяснено, что большинство пикнид гиперпаразита зимой растрескивается и споры гибнут. Поэтому к весне сохраняется лишь небольшой запас инфекции и происходит лишь слабое поражение ржавчины гиперпаразитом. Заметная вспышка развития гиперпаразита наблюдается лишь тогда, когда ржавчинный гриб уже хорошо развился. Поэтому в природе сильное подавление ржавчины за счет естественной инфекции ее гиперпаразитом не происходит. По данным других исследователей, пикниды дарлюки нитчатой хорошо зимуют и выходящие из них конидии могут вызвать сильное поражение ржавчинных грибов. Установлено, что жизнеспособность после перезимовки сохраняют около 85-90% спор гиперпаразита. Отмечено, что зимующий на растительных остатках гиперпаразит в начале лета заражал пустулы стеблевой ржавчины овса и препятствовал дальнейшему ее развитию.
В районах с повышенной влажностью дарлюка нитчатая может снижать поражение растений ржавчиной, развиваясь за счет природного источника инфекции или искусственного распространения спор гиперпаразита.
Два других интересных гиперпаразита ржавчинных грибов - туберкулина кроваво-красная (Tuberculina sanguinea) и туберкулина наибольшая (Т. maxima) - встречаются преимущественно на эцидиях. Эти виды относятся к семейству туберкуляриевых порядка гифомицетов. Первый из этих видов отличается широкой специализацией. Он поражает эцидии, а реже уредо- и телейтопустулы многих видов ржавчинных грибов, образуя в них плоские порошистые коричневато-фиолетовые подушечки. Спороношение гиперпаразита продолжается около 1-1,5 месяца; затем его спородохии становятся твердыми и превращаются в склероции, которые зимуют. Этот вид распространен на ржавчинных грибах в Европе, Азии, Северной и Центральной Америке. В некоторых случаях он может полностью подавлять развитие эцидиев ржавчинных грибов.
Второй вид этого рода - туберкулина наибольшая - узкоспециализированный паразит ржавчинных грибов. Он встречается только на спермогониях и эцидиях видов кронарциум - возбудителей пузырчатой ржавчины сосны в Европе и Северной Америке.
Пораженные спороношения ржавчинного гриба покрываются лиловым или сиреневым налетом конидиального спороношения паразита. Выделения спермогониев стимулируют прорастание спор паразита, и он хорошо на них развивается. Заражает этот гриб только открытые спермогонии и эцидии ржавчинного гриба. Зимует внутритканевый мицелий паразита, а также его с породохии. В некоторых странах этот гиперпаразит при сильном распространении подавляет развитие возбудителя пузырчатой ржавчины. В природе этот гриб является, вероятно, естественным регулятором популяции хозяина. Так, в Северной Америке после применения для борьбы с пузырчатой ржав-чиной сосны некоторых антибиотиков развитие гиперпаразита было подавлено, и это вызвало неожиданную вспышку развития пузырчатой ржавчины.
На различных мучнисторосяных грибах часто паразитирует цицинноболюс Цезати из порядка сферопсидных. Цицинноболюс Цезати, как и дарлюка нитчатая, - космополит, распространенный во всех районах земного шара, где встречается мучнистая роса. Он поражает конидиальное спороношение и клейстотеции хозяина. На конидиальном налете мучнисторосяных грибов при поражении их этим грибом образуются грязновато-серые пятна, постепенно увеличивающиеся в размерах и часто покрывающие весь налет. На поверхности мицелия хозяина, а также в его гифах, конидиях и клейстотециях образуются пикниды гиперпаразита. Они обычно неправильной формы, светло-коричневые или коричневые, с четким сетчатым рисунком. Конидии выделяются из них сильной струей, образуя вокруг пикнид споровую массу. Конидии гиперпаразита переносятся каплями дождя и заражают новые пятна мучнистой росы. При сильном развитии паразита образование конидий и аскоспор у его хозяина полностью подавляется.
Цицинноболюс Цезати можно обнаружить на мучнисторосяных грибах с начала лета и до осени. Особенно обильно он развивается после дождей, при высокой влажности воздуха. Этот гиперпаразит зимует в клейстотециях на листьях, а также в зимующих почках некоторых растений (например, яблони).
При благоприятных для гиперпаразита условиях он может сильно подавлять развитие мучнисторосяных грибов. Нередко он полностью подавляет развитие конидиальной стадии хозяина, а также образование им клейстотециев. Поэтому цицинноболюс Цезати представляет интерес для практического использования его в борьбе с некоторыми мучнисторосяными грибами. В ряде случаев при естественной инфекции и искусственном заражении он сильно снижал поражение яблони мучнистой росой (Д. М. Кобахидзе), а обработка яблонь фунгицидом тиовитом в условиях Ставропольского края не только не снижала развития гиперпаразита, но даже стимулировала его (Г. Р. Дорожио).
В отличие от перечисленных видов, встречающихся в природе только в ассоциации со своими хозяевами, некротрофный микопаразит трихотециум розовый часто обитает как сапрофит на растительных субстратах, вызывает гниль плодов при хранении, а иногда даже паразитирует на ослабленных растениях. Он часто встречается как гиперпаразит на многих фитопатогенных грибах. Этот гриб обнаружен на перитециях гномонии красноустъичной (Gnomonia erythrostoma), стромах полистигмы красной (Polystigma rubrum), нектрии киноварно-красной (Nectria cinnabarina), эпихлое рогозовидной (Epichloe, typhina), склероциях различных грибов, спороношениях возбудителей парши яблони и груши (Venturia inaequalis и V. pirina), ржавчинных и головневых грибов и многих других. Развитие пораженных структур грибов задерживается, или они погибают, и на них образуется ярко-розовый порошистый налет конидиального спороношения паразита.
Способность трихотециума паразитировать на грибах тесно связана с образованием им противогрибного антибиотика трихотецина. Вы-деляемый в среду антибиотик убивает гифы грибов, и трихотециум розовый заселяет их, получая питание из убитых клеток. Штаммы гриба, не образующие трихотецин, не способны развиваться на грибах.
Заселение трихотециумом розовым некоторых обычных его растительных субстратов также связано с его способностью паразитировать на грибах. Например, этот гриб часто вызывает горькую гниль яблок и груш. Однако его мицелий не может проникнуть через неповрежденный эпидермис плодов, и их заражение происходит только при механических повреждениях. Особенно часто этот гриб проникает в плоды через пятна парши. Он заселяет сначала конидиальные спороношения первичного паразита, вытесняет его и развивается затем в ткани плодов. В таких плодах в больших количествах содержится трихотецин, который придает им горький вкус. Накопление грибом антибиотика в ткани плодов предотвращает их заражение другими грибами, и трихотециум розовый развивается в них практически в чистой культуре.
Антибиотик этого микопаразита - трихотецин - подавляет рост многих грибов, в том числе фитопатогенных и возбудителей дерматомикозов. Его получают в промышленных масштабах и используют для защиты растений от некоторых болезней (например, вишни от монилиоза), а также для лечения дерматомикозов у сельскохозяйственных животных и пушных зверей.
Некоторые некротрофные микопаразиты, развивающиеся на плодовых телах дикорастущих или культивируемых съедобных грибов, вызывают их гибель и снижение урожая грибов. В лесах вред, причиняемый микофильными грибами, ничтожно мал по сравнению с тем, который приносят насекомые. Однако в тепличных хозяйствах они могут причинять большой ущерб.
В наших лесах часто можно встретить грибы, покрытые обильным белым мицелием, на котором заметен порошистый налет конидий. Под микроскопом видны мутовчато ветвящиеся конидиеносцы с одноклетными конидиями на вершине веточек. Это широко распространенный во всей умеренной зоне земного шара микофил моноспориум агариковый (Monosporium agaricinum), обитающий на различных гименомицетах, но особенно часто на сыроежках и млечниках (рис. 269). Особенно сильно развивается этот гриб во влажных условиях, после дождей и туманов. На лесных болотах он часто поражает до 60-95% всех сыроежек и млечников, целые группы которых покрывает рыхлый обильный мицелий гриба.
Нередко встречается и другой микофильный гриб - микогоне розовая (Mycogone rosea), образующий хорошо заметный серовато-розовый налет на плодовых телах сыроежек, мухоморов и других агариковых. На его мицелии образуются сначала бесцветные двухклетные конидии на мутовчато разветвленных конидиеносцах, а затем крупные хламидоспоры, состоящие из двух клеток. В массе они имеют розовую окраску. Порошок развивающихся хламидоспор придает характерный вид дерновинкам паразита на плодовых телах грибов. Оба эти микофила вызывают быстрый некроз и разрушение плодовых тел.

Рис. 269. Моноспориум агариковый (Monosporium aga- ricinum) на млечниках
В противоположность этим видам третий микофил, распространенный в умеренном поясе на агариковых,- калъкариспориум арбускула (Calcarisporium arbuscula) - не вызывает быстрой гибели плодовых тел хозяина. Его можно обнаружить в тканях внешне здоровых сыроежек и других грибов. Спороношение у этого микофила развивается на старых плодовых телах в конце лета и осенью. Паразит образует на их шляпках или пластинках темные некротизированные пятна, покрытые очень тонким сероватым налетом спороношения. Хотя этот гриб и образует склероции, их жизнеспособность в почве сохраняется недолго. Мицелий паразита обнаружен в микоризе некоторых сыроежек. Вероятно, это основной способ его перезимовки.
На культивируемом шампиньоне в теплицах известны несколько видов микопаразитов. Один из них - гипомицес розеточный - был описан в разделе, посвященном порядку гипокрейных. Из класса дейтеромицетов на шампиньонах наиболее распространены и вредоносны два вида - вертицилл Молтхауза (Verticillium malthousei) и микогоне губительная (Mycogone perniciosa). Иногда они вызывают эпифитотии.
Если вертицилл Молтхауза поражает молодые плодовые тела шампиньона, вместо них образуются мелкие деформированные клубочки мицелия с сухой поверхностью, покрытой порошистым сероватым налетом конидий паразита, а при заражении крупных, уже сформировавшихся плодовых тел на них образуются коричневые вдавленные некротические пятна, также вскоре покрывающиеся тонким серым налетом спор. Мицелий паразита в засохших плодовых телах может длительное время сохранять жизнеспособность и служить источником инфекции. Сильное развитие вертициллеза шампиньонов происходит при высокой относительной влажности воздуха (до 90-95%) и температуре 15-17 °С. Первичную инфекцию в теплицу могут занести грибные мухи. Нередко она переносится также при сборе плодовых тел грибов. Распространение инфекции в теплице с уже имеющимися очагами инфекции может происходить с каплями воды при поливе.
Другой опасный паразит шампиньонов - микогоне губительная - вызывает у них образование вместо нормальных плодовых тел не дифференцированной бесформенной мицелиальной массы, быстро разрушающейся в результате мокрой гнили. Поверхность таких сильно деформированных плодовых тел покрыта белым войлочным мицелием паразита, на котором, как и у микогоне розовой, образуются два типа спор - конидии и хламидоспоры. Хламидоспоры у этого вида в массе коричневые, с их развитием налет на плодовых телах приобретает коричневую окраску.
Источником первичной инфекции служат хламидоспоры, которые могут сохраняться в почве в жизнеспособном состоянии в течение нескольких лет. Во время плодоношения шампиньонов гриб распространяется конидиями, развивающимися на пораженных плодовых телах.
|
ПОИСК:
|
При использовании материалов проекта активная ссылка обязательна:
http://gribochek.su/ 'Библиотека о грибах'