Порядок Гелоциевые (Helotiales) (А. Г. Райтвийр)
Гелоциевые грибы характеризуются сумками, которые открываются не в виде крышечки на верхушке, как сумки пецицевых грибов, а в виде трещины или поры. Пора обычно довольно хорошо видна в утолщенной стенке верхушки сумки (рис. 121). Плодовые тела гелоциевых в большинстве случаев - типичные апотеции сравнительно малых размеров. Самые маленькие из них не крупнее 0,02 мм в диаметре, а большие достигают лишь 2-3 см в высоту и в диаметре. Плодовые тела еще более крупные встречаются очень редко. Таким образом, в среднем апотеции гелоциевых имеют диаметр 1-3 мм.
Анатомическое строение апотециев у гелоциевых имеет характерные для всех типичных апотециев черты. Однако для апотециев некоторых представителей этого порядка характерно упрощенное строение, вследствие чего в них отсутствует внутренняя ткань - мякоть - между внешним эксципулом (оболочкой апотеции) и субгимением (сплетение гиф под гимением). Такие апотеции состоят только из гимения, тонкого субгимения и гомогенного внешнего эксципул а. Апотеции гелоциевых нередко имеют хорошо развитую тонкую ножку (рис. 111).
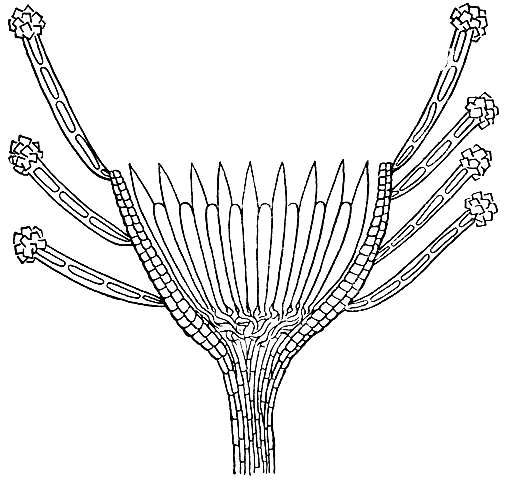
Рис. 111. Строение плодового тела гелоциевого гриба
Споры гелоциевых очень часто асимметрические, многоклеточные, но всегда с гладкой оболочкой. В некоторых семействах встречаются представители с окрашенными спорами.
Гелоциевые грибы - обитатели различных растительных субстратов. Лишь немногие виды можно отнести к напочвенным сапротрофам. Почти полностью отсутствуют представители таких экологических групп, как карбофилы и копрофилы, среди которых можно обнаружить немало пецицевых грибов.
Многие виды гелоциевых грибов активно разлагают субстрат, на котором произрастают. Во время осеннего листопада земля под деревьями в лесу покрывается слоем пожелтевших листьев осины, березы, ольхи, клена. Но еще до того, как ляжет белый зимний покров, от листьев остаются только черешки и жилки. Основную роль в процессе разложения этих листьев сыграл маленький гелоциевый гриб Hymenoscyphus caudatus, плодовые тела которого обильно встречаются в смешанных лесах средней полосы СССР с первой половины сентября до половины октября.
Многие гелоциевые грибы разлагают различные компоненты лесного опада. Важность их роли в круговороте веществ в природе очевидна. Можно подсчитать, что если не было бы грибов, то слой лесного опада увеличивался бы в год примерно на 3-5 см и за 100 лет достиг бы толщины в несколько метров.
Наиболее благоприятным местообитанием гелоциевых грибов являются сообщества высокотравья. Здесь на прошлогодних, отмерших стеблях происходит массовое развитие самых различных гелоциевых грибов.
Большинство гелоциевых грибов является сапротрофами, но во многих семействах встречаются и паразитные виды. Почти все грибы из семейства склеротиниевых (Sclerotiniaсеае) - паразиты.
Во многих случаях паразитический образ жизни связан с увеличением значения конидиальной стадии в цикле развития. У сапротрофных видов конидиальное плодоношение встречается сравнительно редко.
Половой процесс у гелоциевых грибов осуществляется с помощью спермациев, которые выполняют роль мужских половых клеток. Спермации развиваются в особых органах - спермидиях. Иногда их называют и микроконидиями, но, в отличие от настоящих конидий (макроконидий), они не служат средством распространения гриба.
Примером полового процесса у гелоциевых грибов может служить оплодотворение у представителя семейства склеротиниевых Stromatinia gladioli.
Спермидии этого гриба состоят из пучка разветвленных сперматофоров, вырастающих из одной клетки воздушного мицелия. Они похожи на маленькие млечные капельки, диаметром 1,5 мм. Спермации развиваются в больших количествах. Они шаровидные, диаметром 1,2-1,8 мкм, погружены в слизистую жидкость, которая при высыхании приобретает восковидную консистенцию, но легко растворяется в воде.
Женские половые органы у S. gladioli вырастают из стромы в виде столбиков, высотой 0,8-1,9 мм и шириной 0,4-0,8 мм. Они светло-коричневые, покрыты волосками и тонким слоем слизистого вещества. Внутри столбика можно видеть аскогон, состоящий из закрученных гиф с редкими перегородками и многоядерными клетками. От аскогона вырастают к верхушке столбика трихогинные гифы. Оптимальная температура для развития женских половых органов гриба S. Gladioli - + 13° - 24°. Спермации, попавшие на верхушки столбиков, оплодотворяют аскогон, что сопровождается развитием апотециев. Обычно столбик с аскогоном разветвляется после оплодотворения и дает начало нескольким зачаткам апотециев. Апотеции развиваются в течение двух недель после оплодотворения.
S. gladioli - гетероталличный вид: аскогоны не оплодотворяются спермациями с того же мицелия. Оплодотворение может осуществляться спермациями с мицелия другого полового типа.
У других гелоциевых грибов половой процесс осуществляется в основном таким же путем. Различия зависят от того, в какой стадии развития вегетативных органов гриба появляются спермидии и аскогоны.
Обычно это происходит раньше, чем у S.gladioli, аскогоны которого развиваются лишь после того, как уже практически закончился вегетативный рост гриба и образовалась строма, где откладываются питательные вещества, используемые после периода покоя. Наблюдать аскогоны довольно трудно, и поэтому половой процесс описан детально лишь у некоторых видов.
Спермации известны у многих видов из всех семейств гелоциевых грибов, и экспериментально установлено, что среди последних есть и гетероталличные, и гомоталличные виды.
Семейство Геоглоссовые (Geoglossaceae)
Это семейство объединяет немногочисленную группу гелоциевых грибов, характеризующихся сравнительно крупными булавовидными или лопатовидными плодовыми телами. За редким исключением, они почти всегда - напочвенные сапротрофы; плодовые тела их могут достигать 10 см высоты и 2 см в диаметре. Плодовые тела геоглоссовых имеют хорошо развитую ножку, и по строению они являются модифицированными апотециями, у которых выпуклый диск вырос в вытянутую верхнюю часть плодового тела и гимений покрывает наружную поверхность образовавшейся таким образом шляпки (рис. 112).
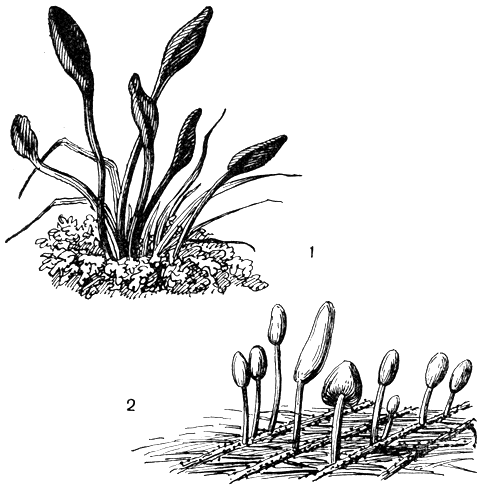
Рис. 112. Гелоциевые: 1 - трихоглоссум волосистый (Trichoglossum hirsutum); 2 - митруля палюдоза (Mitru a paludosa)
У геоглоссовых грибов преобладают длинные цилиндрические многоклеточные споры, которые у представителей родов геоглоссум (Geoglossum) и трихоглоссум (Trichoglossum) окрашены в бурый цвет. Одноклеточные споры встречаются у некоторых грибов рода микроглоссум (Microglossum), у более примитивных представителей семейства спрагуола (Spragueola) и др. Геоглоссовым грибам весьма характерно сильное и разнообразное развитие верхних клеток парафиз. Они часто расширены, имеют различную форму и выступают над сумками, образуя эпитеции.
Обычные представители геоглоссовых грибов в лесах умеренного пояса - спатулария (Spathularia flavida) и кудония (Cudonia circinans), произрастающие на подстилке в ельниках и смешанных хвойных лесах. Нередко они образуют "ведьмины круги", и в одном месте можно найти десятки плодовых тел. Оба вида считают съедобными грибами, но они не имеют практического значения из-за малого размера плодовых тел.
В болотах и на заболоченных лугах нередко можно увидеть черные булавовидные плодовые тела трихоглоссума волосистого (Trichoglossum hirsutum). Этот гриб влаголюбив, как и остальные представители этого семейства. Его плодовое тело покрыто многочисленными темно-бурыми игловидными щетинками, так что весь гриб кажется бархатистым. Споры трихоглоссума волосистого длинные, цилиндрические, шестнадцатиклеточные, окрашенные в бурый цвет.
В сумке они расположены параллельно друг другу, в одном пучке. Из-за этой особенности они не могут освобождаться одновременно или почти одновременно, как споры других дискомицетов, а выбрасываются из сумок в определенной последовательности. Когда сумка созревает, она лопается: в ее верхней части образуется небольшое круглое отверстие. Тотчас же одна из спор выдавливается в открывшуюся пору, полностью закупоривая сумку. Под влиянием гидростатического давления в сумке эта спора выдавливается из нее (рис. 113). Пока не выйдет около половины споры, она двигается очень медленно. Затем спора быстро набирает скорость и в конце концов выбрасывается с такой быстротой, что за ней нельзя уследить до тех пор, пока ее движение не замедлится вследствие сопротивления воздуха и она не промелькнет на расстоянии около 0,5 см от поверхности плодового тела. Немедленно после выбрасывания одной споры ее место занимает другая, закрывающая пору на верхушке сумки прежде, чем сумка подвергнется какому-либо съеживанию. Эта спора, в свою очередь, вылетает через несколько секунд после первой и так далее, пока не будет выброшен весь комплект из восьми спор. После выбрасывания последней споры сумка быстро съеживается, поскольку больше нет спор, которые действовали бы в качестве временной пробки.
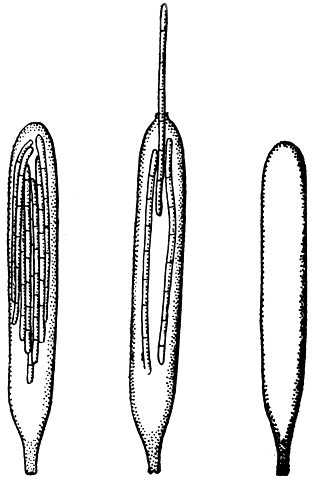
Рис. 113. Выход аскоспор у гелоциевого гриба
Выбрасывание спор у грибов родов трихоглоссум и геоглоссум характеризуется также тем, что оно происходит не "взрывами" в ответ на механический толчок от соприкосновения с апотециями или от движения воздуха, как у большинства дискомицетов, а в виде постепенного процесса, который усиливается под влиянием солнечной радиации. Поэтому указанный тип выбрасывания спор называется радиосенситивным, в отличие от более распространенного тактиосенситивного ("взрывами", "залпами").
Многие виды геоглоссовых - гигрофилы или умеренные гигрофилы. Обычно их можно найти во влажных местообитаниях. Эти грибы также сравнительно теплолюбивы, и ареал многих видов совпадает с распространением широколиственных пород, хотя они и предпочитают смешанные и хвойные леса. На Дальнем Востоке встречается самая богатая в Советском Союзе флора геоглоссовых. В сезон муссонных дождей в хвойно-широколиственных лесах Хабаровского края и Приморья почти на каждом шагу встречаются черные палочки геоглоссума и трихоглоссума. Видовое разнообразие их здесь также большое.
Наряду с ними на Дальнем Востоке растут и Microglossum fumosum оливкового и Microglossum rufum оранжевого цвета. Эти виды распространены и в Северной Америке. Они свидетельствуют о тесных связях между грибной флорой Северной Америки и Восточной Азии.
Семейство Склеротиниевые (Sclebotiniaceae)
Наиболее характерный признак всех грибов этого семейства - наличие склероция или стромы, из которых вырастают апотеции. Подавляющее большинство видов этого семейства - паразиты. Образование их склероциев связано с особенностями их жизненных циклов.
Апотеции склеротиниевых грибов развиваются, как правило, весной. Исключение составляют сапротрофные виды рода рутстроемия (Butstroemia), которые причисляются к осенним грибам. Из почвы появляются крупные, более 1 см в диаметре, коричневые бокальчики Sclerotinia tuberosa (табл. 21). Их можно всегда найти только в растительном сообществе с ветреницами. Если выкопать ножку гриба, то оказывается, что она начинается из крупного шаровидного черного склероция на корневище ветреницы. Другие склеротиниевые грибы можно встретить на прошлогодних мумифицированных цветках или плодах разных растений: на сережках осины, березы, ольхи, на плодах березы и осоки, на ягодах брусники, голубики, клюквы, на диких яблоках и грушах. Встречаются они на стеблях тростника и осоки, а также на многих субстратах.

Таблица 21. Гелоциевые грибы: вверху слева направо: склеротиния клубневая (Sclerotinia tuberose), калицелла цитрусовая (Calicella citrina), дазисцифелла калициевидная (Dasiscyphella caliciformis); внизу слева - монилиния фруктовая (Monilinia frutigena); внизу справа - корине саркоидная (Coryne sarcoides)
Отмечено, что склеротиниевые грибы плодоносят одновременно с цветением их растений-хозяев. Это тесно связано с биологией распространения склеротиниевых грибов.
Апотеции склеротиниевых грибов обладают позитивным фототропизмом, в результате которого они всегда стремятся к свету. Благодаря этому апотеции вырастают из склероциев, которые часто погружены в почву или в слой прошлогоднего опада, к открытому воздуху и выбрасывают аскоспоры так, что они захватываются и разносятся ветром. Аскоспоры выбрасываются из сумок силой осмотического давления, когда лопаются поры в утолщенных верхушках сумок. Это случается, если, например, ветер сотрясает апотеции, что служит толчком для одновременного открывания всех зрелых сумок в апотеции.
Все зрелые споры разбрасываются одним "залпом" на довольно большое расстояние от плодового тела (например, около 5 см у Sclerotinia tuberosa). Склеротиниевые грибы по типу освобождения спор относятся к типично тактиосенситивным дискомицетам.
Споры, попавшие на подходящий субстрат, прорастают, но первичный мицелий обладает очень слабой жизнеспособностью и малой способностью к проникновению в растительные ткани. Это объясняется тем, что аскоспоры склеротиниевых грибов содержат мало питательных веществ. Удачная инфекция обеспечивается лишь в том случае, когда в месте попадания споры имеются определенные питательные вещества и стимуляторы роста, которые встречаются в ранах, в выделениях субкутикулярных тканей или в железистых выделениях рыльцевых клеток. Поэтому лучше всего прорастают те споры склеротиниевых грибов, которые попадают на рыльце их растений-хозяев. Мицелий от проросшей на рыльце споры легко проникает через канал пестика во внутренние части цветка и оттуда - в вегетативные части растения. У многих видов склеротиниевых грибов заражение происходит исключительно через цветки растений-хозяев, чем и объясняется совпадение времени плодоношения гриба со временем цветения растения-хозяина. Некоторые склеротиниевые грибы заражают своего растения-хозяина и через молодые, только что распустившиеся листья, тогда время плодоношения гриба совпадает со временем облиствления.
После удачного заражения склеротиниевые грибы питаются за счет своих растений-хозяев. К настоящим паразитам, которые получают питательные вещества из живых клеток своего хозяина, не убивая их, относятся лишь немногие виды склеротиниевых. Грибы большинства видов относятся к некрогенам и некрогрофам, гифы которых сперва убивают клетки и ткани хозяина, а затем питаются уже мертвым органическим веществом. Даже грибы паразитарных видов в начальных стадиях развития являются паразитами и лишь позднее превращаются в некротрофы.
Некоторые грибы (например, Sclerotinia sclerotiorum) проникают при благоприятных условиях в любые части растения-хозяина и уничтожают его, но они бывают скорее исключениями среди склеротиниевых грибов. Большинство видов этого семейства развивается в строго ограниченных частях хозяина: или в стеблях, или в корневищах, листьях, сережках, плодах, где и образуются склероции. Склероции - органы накопления и сохранения пищи. Они у различных представителей склеротиниевых грибов бывают разного типа.
У грибов рода склеротиния (Sclerotinia) развиваются клубневидные склероции, свободно образующиеся на воздушных гифах. Их форма зависит от формы тех полостей в органах растения-хозяина, где они развиваются. Если они развиваются без влияния внешнего давления, они имеют шаровидную форму. В строении склероциев можно различать черную кору и белую (или сероватую) мякоть. Все запасы питательных веществ содержатся в толстых гифах мякоти. Когда из склероциев вырастают апотеции, эти питательные ресурсы используются полностью и склероции превратятся в пустые шарики.
Полые шаровидные склероции характерны для грибов рода монилиния (Monilinia). Они образуются под эпидермисом пораженного плода и покрыты снаружи и изнутри черной корой.
Мумиевидные склероции, характерные для грибов рода цибория (Ciboria), подражают форме пораженных органов - сережкам или семенам. Они образуются путем переваривания тканей субстрата и замещения их гифами мякоти.
Типичные склероции у грибов рода рутстроемия (Butstroemia) отсутствуют, вместо них развиваются субстратные стромы. Такая строма видна на поверхности поврежденного листа в виде черной корочки. Эта корочка - кора стромы, которая окружает те ткани субстрата, которые гриб "запасает" для питания. Внутри стромы ткань субстрата пронизана рыхлой сетью сплетения тонких гиф гриба. Примерно в то же время, когда начинают развиваться склероции, появляются и спермации.
Бесполое размножение склеротиниевых грибов осуществляется с помощью конидий, но они встречаются далеко не у всех представителей этого семейства. Конидиальное плодоношение наиболее характерно для представителей родов монилиния и ботриотиния.
Многие склеротиниевые грибы - важные возбудители болезней растений.
Грибы этого рода образуют склероции в плодах разных растений из семейств розоцветных и брусничных. В цикле развития у них всегда встречается конидиальное плодоношение типа монилия (Monilia). Многие грибы этого рода были впервые подробно изучены крупным русским микологом М. С. Ворониным.
Рассмотрим цикл развития гриба Monilinia urnula, паразитирующего на бруснике. Апотеции М. urnula развиваются из склероциев примерно в середине или во второй половине мая. Аскоспоры попадают на молодые побеги брусники и прорастают. Ростковые трубки проникают через клетки эпидермиса в стебли и листья растения, вызывая характерное заболевание. Стебель пораженного молодого побега становится на определенном протяжении вялым, сморщивается и засыхает (при этом стебель вначале желтеет, затем буреет и наконец почти чернеет), в большинстве случаев он изгибается. Листья на пораженном участке стебля также буреют, чернеют и засыхают, но чаще всего засыхает и чернеет только нижняя поверхность пластинки листа, а верхняя остается свежей, зеленой.
Гифы гриба распространяются по стеблю межклеточно, вначале в камбиальном слое, затем в древесине и даже в сердцевине. В коре их в это время еще нет, хотя кора по большей части уже отмирает и окрашивается в бурый цвет. Гриб выделяет вещества, убивающие клетки стебля брусники, и питается уже мертвыми частями пораженного стебля, являясь, таким образом, типичным некротрофом.
После того как в пораженной части стебля все слои коры убиты, начинается бурное и пышное дальнейшее развитие гриба. Теперь появляются в очень большом количестве гифы, которые распространяются между клетками внутренней паренхимы коры и растут в сторону ее наружной части. Чем больше гифы приближаются к наружной мелкоклеточной зоне коры, тем толще они становятся. Достигнув этой зоны, они распространяются в ней как в радиальном, так и в вертикальном направлении, между отмершими клетками коры и эпидермиса, а также между эпидермисом и кутикулой. В гифах образуется много перегородок, и клетки приобретают более или менее округлую или эллипсоидальную форму. Таким образом, в наружной зоне коры образуется бесцветная псевдопаренхиматическая грибная строма. Она развивается чаще всего на одной стороне стебля, что и вызывает коленообразный изгиб пораженного побега.
Из стромы вырастают гифы, которые разрывают кутикулу. Выходящие из стромы гифы всегда снабжены расположенными на одинаковых расстояниях перетяжками, которые придают им характерную четковидную форму. Вначале эти гифы не имеют ни одной перегородки. Вскоре наступает момент, когда дальнейший верхушечный рост гиф прекращается и они распадаются на отдельные клетки. Каждая клетка теперь становится конидией.
Масса конидий появляется на стебле или на нижней поверхности листьев в виде довольно плотного снежно-белого или слегка желтоватого налета. Теперь пораженные, покрытые конидиями стебли издают сильный, очень приятный, миндальный запах. Этот запах привлекает пчел и мух, которые посещают больное растение и переносят конидии на рыльца расцветающих к этому времени цветков здоровых растений.
Попадая на рыльце, конидии тотчас же прорастают в них в виде длинных ростковых трубочек, которые по каналу пестика проникают в гнезда молодой завязи брусники, постепенно заполняющиеся гифами гриба. В этот период развития гриба зараженные ягоды не отличаются от здоровых. Но в дальнейшем гифы гриба проникают в стенки плода и начинается образование склероциев полого шаровидного типа. Затем, примерно в то же время, когда здоровые ягоды начинают краснеть, пораженные приобретают грязную желтовато-бурую окраску, затем темнеющую и становящуюся каштаново-бурой.
Одновременно тонкая кожица ягоды сохнет, сморщивается и плотно прижимается к склероцию. Очертания четырех дугообразных ребер склероциев очень отчетливо выступают через кожицу, и каждая ягода, содержащая склероции, становится сплющенной и ребристой.
Ягоды со склероциями очень легко отделяются от плодоножек и перезимовывают на мху, среди лесного опада. Весной, когда снег тает, в склероциях, еще под снежным покровом при сравнительно низкой температуре, образуются зачатки плодовых тел. Апотеции вырастают по одному из каждого склероция и созревают в конце апреля или начале мая.
Monilinia ledi образует склероции в плодах багульника. Этот гриб примечателен тем, что у него встречается резко выраженная разнохозяйность. Конидиальную стадию его нельзя найти на багульнике, так как она встречается на молодых побегах голубики. Такая смена питающих растений, обычная для ржавчинных грибов, в других группах грибов встречается очень редко, и М. ledi - интересный пример этого феномена.
Monilinia fructigena является наиболее известным и важным в хозяйственном отношении видом этого рода. Этот возбудитель черной гнили яблок и других семечковых плодовых культур встречается практически в каждом фруктовом саду. Пораженные грибом яблоки буреют и покрываются желтоватыми подушечками конидиеносцев (табл. 21). Первичное заражение происходит посредством конидий, через рыльце, но конидии с больных яблок заражают все новые и новые плоды. В пораженных плодах развиваются полые шаровидные склероции, в результате чего яблоки чернеют.
В отличие от других грибов рода Monilinia у M. fructigena сумчатая стадия (апотеции) развивается исключительно редко. Чаще всего перезимовывающие чсклероции прорастают весной снова в виде конидий. Так, у M. fructigena и возбудителя гнили косточковых пород M. cinerea наблюдается сильное уменьшение значения сумчатой стадии в цикле развития. Известны лишь немногие случаи, когда при очень благоприятных условиях отмечено развитие апотециев M. fructigena.
Виды этого рода растут преимущественно на представителях семейства осоковых. Весной, во время цветения, аскоспоры, переносимые ветром, попадают на рыльце и прорастают там; Зараженные соцветия погибают и буреют после проникновения гриба в отдельные цветки. Зараженные стебли никогда не плодоносят. Гифы гриба растут медленно вниз по стеблю и под его эпидермисом образуют спермации. Они прорываются через эпидермис и появляются на стебле в виде изумрудных или коричневатых слизистых капель. Дождь, роса и насекомые распространяют спермации по умирающему стеблю вниз, где начинают развиваться склероции. Спермации участвуют в процессе оплодотворения в качестве мужских половых клеток, но точный механизм оплодотворения пока не изучен. Конидиальное спороношение у видов рода Sclerotinia отсутствует и склероции развиваются в стеблях растений-хозяев. Сначала они напоминают кусочки ваты, но постепенно твердеют, у них дифференцируются черная кора и розоватая мякоть, позднее становящаяся белой. Поздно осенью, когда склероции созревают, они выступают из разложившихся стеблей. У некоторых грибов они выпадают в воду или на моховой покров вокруг растений-хозяев. У других они остаются внутри стеблей, и выступает только частично их поверхность. Гриб каждого вида имеет склероции более или менее характерной формы, которая при этом очень во многом зависит от условий развития гриба в стеблях. Склероции перезимовывают среди растительных остатков, и следующей весной из них вырастают апотеции.
Многие виды рода склеротиния строго специализированы к определенным питающим растениям, в частности те виды, грибы которых развиваются на растениях семейства осоковых. Но есть в роде склеротиния вид с очень широким физиологическим спектром, поражающий широкий круг питающих растений из разных семейств. Это - S. sclerotiorum.
S. sclerotiorum хорошо известен как возбудитель белой гнили различных сельскохозяйственных культур. Он встречается на корнях свеклы, цикория, моркови, петрушки, на стеблях подсолнечника, томата, фасоли, бобов, гороха, салата, льна и многих других, на плодах огурцов и кабачков, томатов, гороха, фасоли, бобов, на соцветиях сложноцветных (в частности, подсолнечника), на клубнях картофеля, на кочанах капусты (при хранении) и т. д.
Развитие гриба сопровождается характерными внешними признаками. Поверхность растения покрывается относительно плотным, войлочным, чисто-белым воздушным мицелием гриба, на котором через несколько дней начинают образовываться склероции в виде более плотных и вначале также чисто-белых мицельных скоплений 3-5 мм в диаметре. Зачатки склероциев постепенно сереют, а впоследствии чернеют. В процессе созревания они выделяют много воды в виде бесцветных капелек, которыми склероции как бы усыпаны при развитии во влажной атмосфере. Склероции образуются обычно в больших количествах, группами или рассеянно. Тем временем пораженная ткань растения размягчается и разрушается. Склероции бывают обычно шаровидные или продолговатые, от 0,5 до 4 см в диаметре, но иногда значительно крупнее, имеют форму полостей в тех частях растений, где развивались. Так, в корзинках подсолнечника образуются склероции, заполняющие всю корзинку и имеющие сетчатый вид. Конидиальное спороношение у S. sclerotiorum, как и у других видов рода склеротиния, отсутствует.
Этот гриб встречается довольно часто не только в природе, но и в овощных хранилищах, где является опасным Возбудителем гнили. Гниль распространяется в хранилищах очень быстро, переходя с одного корня на окружающие.
Апотеции развиваются из склероциев по одному (в природных условиях в конце мая - начале июня).
Виды этого рода - паразиты цветков и плодов различных растений. Они отличаются от видов рода монилиния отсутствием конидиальной стадии и строением склероциев. Склероции представителей рода цибория мумиевидные. При поверхностном осмотре они очень сходны с непораженными прошлогодними сережками или плодами питающего растения, но в действительности сережка или плод мумифицированы гифами гриба и превращены в склероции, точно имитирующие внешнюю форму пораженного органа питающего растения. Склероции цибориевых грибов можно отличить от непораженных прошлогодних сережек или плодов только по тому, что из них весной вырастают апотеции, или же при микроскопическом исследовании.
Апотеции грибов Ciboria довольно часто встречаются ранней весной. Цибория - один из самых ранних весенних грибов. На сережках ивы можно найти С. caucus, на сережках ольхи и осины - С. amentacea, на плодах березы - С. betulae. Bpejvm плодоношения всех их совпадает со временем цветения питающего их растения.
Такие виды, как С. caucus и С. amentacea, обитают только в мужских сережках питающих растений. Ко времени распускания сережек апотеции грибов уже вполне развиты и выбрасывают зрелые аскоспоры. Им, как и многим другим представителям склеротиниевых грибов, характерна способность расти и развиваться при низкой температуре. Попавшие на тычинки аскоспоры сразу прорастают, и ростовые трубки проникают в тычинки. Мицелий гриба очень быстро захватывает всю сережку. Зараженные сережки быстро вянут, а затем мумифицируются гифами гриба. Склероции состоят из тесно сплетенных гиф, среди которых можно найти остатки клеток питающего растения. Опавшие на землю склероции перезимовывают, на них развиваются спермидии и аскогонии и после оплодотворения вырастают апотеции.
Семейство Гиалосцифовые (Hyaloscyphaceae)
К семейству гиалосцифовых относятся те гелоциевые грибы, плодовые тела которых покрыты волосками. Их изящные бокальчики и чашечки встречаются очень часто на отмерших стеблях растений, на опавших листьях, на гниющей древесине. С помощью лупы и микроскопа можно увидеть богатство форм гиалосцифовых грибов. Одни из них покрыты редкими прямыми, другие - густыми извилистыми волосками - белыми, ярко-желтыми, бурыми, фиолетовыми, красными. На их верхушках можно увидеть разнообразные скопления кристаллов.
Волоски, которые покрывают снаружи апотеции, - характерный признак всего семейства, но различия в форме, цвете и строении волосков служат основой систематики этого семейства. При этом волоски служат не просто покрывалом для апотециев, а играют важную функциональную роль в жизни гриба. У многих видов волоски толстостенные и с толстыми перегородками, а в клетках волосков всегда содержится живая цитоплазма и живые ядра. Волоски действуют как секреторные органы и выделяют в виде оксалата кальция или различных смолистых веществ остатки обмена веществ гриба. Так, различия в строении и инкрустации волосков у гиалосцифовых грибов являются не произвольным систематическим признаком, а указывают на важные различия в физиологии отдельных видов.
Подавляющее большинство гиалосцифовых грибов являются сапротрофами. Паразитизм среди них - явление исключительное, но эти исключения имеют определенное значение в хозяйственном отношении.
Конидиальное спороношение в цикле развития гиалосцифовых грибов не обнаружено. У многих видов известны спермидии и спермации, половой процесс происходит у них по общей схеме.
Сезонному развитию гиалосцифовых грибов характерно появление плодовых тел в первой половине вегетационного периода. У отдельных грибов (например, Belonidium leucophaeum и В. mollissimum) плодовые тела появляются на отмерших стеблях зонтичных уже весной, в начале мая, еще до развития густого травяного покрова. При сухой погоде они остаются незащищенными от солнечных лучей и высыхают. Но все-таки они остаются жизнеспособными и при возобновлении достаточно благоприятных влажных условий вновь оживают и продолжают расти. Оптимум развития плодовых тел наступает после того, как травянистый покров станет достаточно густым, чтобы защищать почву от солнечных лучей и интенсивного испарения воды. Тогда прошлогодние стебли густо покрываются апотециями дазисцифуса (Dasyscyphus), белонидиума (Belonidium) и трихопецицеллы (Trichopezizella). В конце лета и осенью сильно прогнившие стебли пустеют от апотециев, и в это время года наступает сезон плодоношения тех гиалосцифовых грибов, которые обитают на древесине. В сентябре и октябре на коре осины можно обнаружить коричневые чашечки Belonidium corticale, а на сухих обнаженных стволах и ветках лиственных пород - красные апотеции Perrotia flammea.
Род дазисцифус - самый обширный в этом семействе, виды которого распространены как в умеренной зоне, так и в тропиках.
Апотеции дазисцифуса имеют хорошо развитую ножку и покрыты шероховатыми волосками. Они обитают на самых различных субстратах: древесине, опавших листьях и хвое, на стеблях двудольных травянистых растений, злаков, осок. Все виды являются сапротрофами. Самый обычный представитель этого рода - D. bicolor - растет на гниющих стеблях малины. В июне и июле прошлогодние отмершие стебли малины густо покрыты его бокальчатыми снежно-белыми апотециями с ярко-оранжевым гимением. Почти всегда рядом с ними растет и другой вид этого рода - D. clandestinus. У него апотеции коричневые и снаружи покрыты как бы серым налетом. Такой вид придают апотециям накопления кристаллов оксалата кальция на верхушках волосков. Так, гиалосцифовые грибы (как и другие гелоциевые грибы) на определенных субстратах образуют сообщества из нескольких видов.
В этом роде около 20 видов. Эти грибы произрастают на хвойных деревьях. Все они имеют весьма одинаковый облик: апотеции средней величины (1-3 мм в диаметре) с ярко-оранжевым гимением, снаружи снежно-белые. Лишь четыре вида имеют коричневую наружную сторону и покрыты коричневыми же волосками. Некоторые виды этого рода паразитируют на живых стволах и ветках хвойных деревьев и вызывают их заболевания.
Наиболее известный и важный среди паразитных видов рода лахнеллулы - гриб Lachnellula willkommii, вызывающий рак ствола лиственницы. Он поражает чаще всего молодые лиственницы (в возрасте 10-25 лет). Распро-страняется гриб исключительно аскоспорами, которые, попадая на маленькие раны (вследствие мороза, с помощью насекомых и т. п.), прорастают и заражают живые ткани дерева. В результате жизнедеятельности гриба погибает камбий, за чем следует развитие рака на стволе. На краях ракового образования появляются апотеции гриба. Их оранжевый гимений вполне открыт лишь во влажную погоду. В сухую погоду края апотеция загибаются и апотеций как будто закрывается.
Таким образом обеспечивается выбрасывание спор при наиболее оптимальных для прорастания условиях погоды. Это явление связано с высокой гигроскопичностью апотециев и свойственно всем гиалосцифовым грибам, а также многим другим представителям порядка гелоциевых.
При сильном заражении дерева не только появляется рак на стволе, но и засыхают ветви в результате того, что гриб убивает камбий.
L. willkommii широко распространен по всей Европе и причиняет много вреда в северных районах, где патогенное действие гриба комбинируется с повреждениями от мороза, и эти два фактора усиливают друг друга.
На сосне рак вызывает гриб L. pini, имеющий коричневые апотеции с оранжевым гимением. В отличие от L. willkommii этот вид не так широко распространен. Он встречается отдельными очагами в Северной Европе, Карелии, на юге Средней Сибири и на Дальнем Востоке.
Из сапрофитных видов рода лахнеллула (Lachnellula) очень часто встречается гриб L. suecica, который можно обнаружить на сухих веточках всех хвойных пород. Он растет и на юге, и на самом далеком севере: его можно найти даже на сухих веточках одиноких лиственниц в лесотундре. Очень типичен и L. calyciformis, который поселяется на коре сваленных стволов пихты.
Представители этого рода отличаются сидячими, нередко ярко окрашенными апотециями.
В отличие от представителей предыдущих родов белонидиум имеет волоски с гладкими стенками. Встречаются они обильно на гниющих стеблях травянистых растений. Уже рано весной появляются обросший ярко-желтыми волосками Belonidium leucophureum и снежно-белый В. mollissimum. Они обильны еще в первой половине лета, но затем заменяются видами В. sulphureum и В. adenostylidis, которые по внешнему облику вполне напоминают свои виды-двойники, но отличаются от них микроскопическим строением. Некоторые виды рода имеют характерно альпийское или аркто-альпийское распространение: они встречаются только в высокогорьях или же в высокогорьях и Арктике. Такие виды имеют более интенсивную пигментацию, что защищает их от интенсивного солнечного излучения высокогорья и в условиях длинного полярного дня. На северной границе леса и в высокогорье Тянь-Шаня нередко встречается В. elegantulum, который имеет апотеции с тускло-красным гимением и темно-лилово-бурой наружной стороной.
Семейство Гелоциевые (Helotiaceae)
Это самое обширное и разнообразное семейство порядка. Его представители принадлежат к различным жизненным формам. Здесь можно найти грибы с нежными гигроскопическими апотециями, и с плодовыми телами типа шляпочных грибов, и крупные студенистые апотеции, которые представляют резервуары воды.
Общая черта всего семейства - анатомическое строение апотециев: между субгимением и внешней корой имеется хорошо развитый слой мякоти из рыхло переплетенных гиф, а внешняя кора состоит из более или менее призматических клеток. Конидиальная стадия известна у немногих видов. У некоторых видов конидиальное спороношение непосредственно предшествует развитию апотециев или же конидиальные стромы встречаются рядом с апотециями. Большинство видов - сапротрофы на отмерших частях растений, но встречаются и паразиты.
Наиболее типичными представителями семейства гелоциевых можно считать обширные роды гименосцифус (Hymenoscyphus) и фиалея (Phialea). Грибы обоих родов имеют светлоокрашенные апотеции на длинных ножках, некоторые из них встречаются в массовом количестве.
Среди этих грибов упомянутый выше Hymenoscyphys caudatus разлагает опавшие листья. Также обильно встречается на отмерших стеблях очень многих травянистых растений Phialea cyathoidea. Его бокальчатые апотеции на длинных тонких ножках густо покрывают прошлогодние стебли зонтичных, крапивы, таволги с весны до конца лета. Время плодоношения P. cyathoidea совпадает со временем плодоношения травообитающих гиалосцифовых грибов. Они вместе характеризуют весенне-летний аспект обитающих на травах грибов. Их плодоношение приходит к концу во второй половине лета, и их место занимает осенний аспект, который характеризуется массовым развитием гименосцифусов - Hymenoscyphus scutula, Н. herbarum и других видов этого рода.
Поселяющиеся на деревьях гименосцифусы, например Н. salicellum, Н. calyculus, принадлежат даже к позднеосеннему аспекту. Их апотеции растут на опавших ветках различных лиственных пород от сентября до ноября, практически до выпадения снега. Температура до -10° С не повреждает их плодовых тел. Часто бросаются в глаза маленькие ярко-желтые чашечки, которые тесными группами растут на сваленных стволах. Это калицелла (Calycella citrina, табл. 21) - один из самых обыкновенных представителей семейства в смешанных лесах средней полосы СССР.
В этом же семействе встречается целая группа представителей со студенистыми плодовыми телами. Самый характерный среди них гриб - булгария (Bulgaria inquinans). Он имеет сравнительно крупные, диаметром до 3 см, черные апотеции, которые группами вырастают из-под коры опавших стволов дуба или (реже) других лиственных пород. Снаружи они покрыты складчатой кожистой корой, под которой скрывается студенистая, очень богатая водой мякоть. Апотеций В. inquinans представляет таким образом как бы резервуар воды, который необходим для его жизнедеятельности и споруляции. И действительно, оторванные от субстрата апотеции продолжают еще сравнительно долго выбрасывать черные аскоспоры, которые покрывают все вокруг гриба, как налетом сажи.
Нередко встречается на гниющих пнях и другой гриб со студенистыми апотециями Корине саркоидная (Coryne sarcoides, табл. 21). Его апотеции мясисто-красные или фиолетовые. Перед их развитием на том же месте растет конидиальное плодоношение этого гриба. Конидиальные стромы лопатчатые, студенистые, такой же окраски, как и апотеции. Конидии развиваются на коротких бесцветных конидиеносцах, которые покрывают верхнюю часть стромы слизистой оболочкой. Переносятся они водой (дождем и росой).
Немногие представители этого семейства принадлежат к шляпочным грибам. Их апотеции имеют хорошо развитую ножку и булавовидную шляпку, приросшую нижним краем к ножке. Самый крупный среди таких шляпочных гелоциевых грибов - митруля палюдоза (Mitrula paludosa). Плодовые тела этого гриба бывают высотой до 3 см, имеют снежно-белую ножку и ярко-оранжевую шляпку. М. paludosa отличается ярко выраженной гидрофильностью и растет на опавших в воду гниющих листьях. В средней полосе СССР он относится к весенним грибам, так как оптимум развития его плодовых тел находится при невысокой температуре. В южных районах страны его можно найти в высокогорьях в течение всего лета. Там М. paludosa растет в узких тенистых каньонах маленьких высокогорных ручейков прямо в воде или там, где почва регулярно опрыскивается водой с температурой 8-12°С (рис. 112).
Сравнительно обособленную позицию в семействе занимают представители подсемейства энцелиоидовых (Encoelioidea). Их плодовые тела растут тесными группами и прорываются пучками через кору субстрата. Отдельные апотеции сравнительно крупные, в среднем около 1 см в диаметре. Снаружи они покрыты характерным мучнистым налетом. Сюда относят один из самых ранних грибов - энцелию фасцикулярис (Encoelia fascicularis), апотеции которого прорываются пучками через кору опавших стволов осины (рис. 114). Группы коричневых апотециев контрастно выступают на светлосерой коре осины. На ольхе растет близкий к предыдущему вид Е. furfuracea. В странах Западной Европы, где климат более мягкий, оба вида относятся к зимним грибам: там сезон их плодоношения продолжается от декабря до мая, когда стоит благоприятная для этих грибов прохладная и влажная погода.

Рис. 114. Энцелия фасцикулярис (Encoelia fascicularis)
Семейство Дермateацевые (Dermateaceae)
К этому семейству принадлежат те гелоциевые грибы, в апотециях которых отсутствуют дифференцированные мякоть и кора. Их апотеции построены главным образом из мясистой ткани, которая состоит из округлых клеток с бурыми тонкими стенками, и, как правило, не имеют ножку. Многие представители имеют очень маленькие и незаметные апотеции. Несовершенная стадия встречается чаще, чем в других семействах гелоциевых грибов. Среди представителей дерматеацевых грибов встречаются сапротрофные и паразитические виды.
Типичным сапротрофным представителем этого семейства служит гриб из рода моллизия (Mollisia cinerea). Его апотеции сидячие, дисковидные, маленькие, до 3 мм в диаметре, но хорошо заметные, потому что растут скученно и их светло-серые диски отчетливо выступают на темной поверхности мертвой коры или древесины. М. cinerea появляется уже весной, и его можно найти до поздней осени. Очень похожи на грибы рода моллизия представители рода тапезия (Tapesia). Однако они отличаются тем, что их апотеции образуются на хорошо развитом субикулюме из темно-бурых или черных гиф.
Грибы родов псевдопецица (Pseudopeziza) и лептотрохила (Leptotrochila) ведут паразитический образ жизни. Они растут на живых листьях различных цветковых растений, вызывая пятнистость листьев. На листьях люцерны растет Leptotrochila medicaginis. Он вызывает коричневатые пятна на листьях люцерны. По краям пятна расположены группы конидиеносцев, на которых образуются булавовидные конидии. Маленькие (в среднем диаметром 0,5 мм) чернью апотеции развиваются в центрах пятен. Оптимальная температура для роста этого гриба +20° С. Он обладает и некоторыми ксерофильными чертами: оптимальной влажностью воздуха для освобождения спор из сумок является 98%. Освобождение спор происходит так же, как и у других дискомицетов: они выбрасываются из сумок силой осмотического давления, что должно быть не меньше 5,7 атм. Такое давление дает спорам начальную скорость 32,4 м в 1 сек и выбрасывает их на расстояние до 40 мм от апотеция. Необходимым условием для споруляции является максимальный тургор апотеция.
В отличие от многих других дискомицетов L. medicaginis спорулирует по очень строгому ритму. Освобождение спор происходит каждый день от 6 до 10 ч, причем в один день выбрасывается около 2000 спор. Половина из этой ежедневной нормы может выбрасываться всего за 10 мин. Один апотеций продолжает спорулироваться 8-14 дней и может при этом продуцировать до 30 000 спор.
|
ПОИСК:
|
При использовании материалов проекта активная ссылка обязательна:
http://gribochek.su/ 'Библиотека о грибах'