Порядок Мукоровые (Mucorales) (А. А. Милько)
Мукоровые грибы занимают довольно обособленное положение среди остальных низших грибов. Обнаруживая немало общих черт в строении таллома (гифальная структура) с оомицетами (сапролегниевые, пероноспоровые), мукоровые одновременно с этим отличаются от них целым рядом морфолого-биологических особенностей.
Таллом мукоровых грибов состоит из хорошо развитых ветвящихся гиф. Гифы обычно неокрашенные, и длина их колеблется от нескольких миллиметров до нескольких сантиметров. Молодые (развивающиеся) гифы одноклеточные, и только у немногих представителей этого порядка (семейство кикселловых) они многоклеточные, чем напоминают таковые сумчатых и других, более высокоорганизованных форм.
У большинства мукоровых грибов поперечные перегородки в гифах образуются при старении культуры или только в местах локализации репродуктивных органов.
У мукоровых, как и большинства остальных грибов, гифы довольно однообразны в морфологическом отношении, в связи с чем они не имеют систематического значения. Такое значение для отдельных групп и видов порядка имеют их видоизменения, к которым относятся столоны, ризоиды, хламидоспоры, оидии, аппрессории, гаустории и т. п.
У большинства мукоровых грибов известны два способа размножения (спороношения) - бесполое и половое. Однако оба эти способа размножения неравноценны по своему значению в распространении и сохранении в природе грибов данного порядка.
Мукоровые грибы, за небольшим исключением (некоторые виды рода зигоринхус), активно развивают бесполое спороношение, что определяет первостепенное значение его в распространении и сохранении вида в природе. Развитие этого спороношения при благоприятных условиях происходит равномерно на протяжении всего онтогенеза гриба. Исключение в данном случае составляют представители рода пилоболус, у которых спороношение развивается с угасающей ритмичностью и только на ранних стадиях онтогенеза гриба.
Мукоровые грибы характеризуются различной сложностью строения аппарата бесполого размножения, но основными его элементами являются спороносцы и споры.
Спороносцы представляют собой морфологически дифференцированные боковые ответвления вегетативных гиф (табл. 9). Они бывают нитевидные, цилиндрические, шиловидные или булавовидные, простые или различно разветвленные, что имеет важное видовое и родовое систематическое значение. Возникают спороносцы в виде почковидного выроста морфологически недифференцированных участков гиф или их видоизменений (столона, ризоида), нарастающего верхушкой. Исключение составляют грибы родов пилоболус и утаромицес, у которых спороносцы развиваются из особых клеток гиф субстратного мицелия, именуемых трофоцистами.
У многих мукоровых грибов спороносцы в период своего активного роста обладают положительным фототропизмом, т. е. изгибаются по направлению к источнику света. Такой особенностью характеризуются, как правило, простые спороносцы (стилоспорангиеносцы) и преимущественно у грибов родов фикомицес, пилаира, пилоболус и у крупных форм рода мукор. Предполагается, что β-каротин, флавин или рибофлавин являются теми веществами содержимого спороносцев, которые, поглощая свет определенной длины волны и действуя как раздражитель, обусловливают их положительный фототропизм.
Споры бесполого размножения (спорангиоспоры, мероспорангиоспоры, конидии) мукоровых грибов представляют собой одноклеточные неподвижные образования. Они имеют различную форму, что важно для определения видов. Для прорастания споры не нуждаются в предварительном периоде покоя. Они прорастают вегетативно ростковой трубкой, которая быстро разрастается, образуя мицелий. Распространяются споры воздушным течением, человеком и животными.
При бесполом размножении споры возникают эндогенно (в споровместилищах) или экзогенно (на вздутиях спороносцев). В зависимости от способа образования этих спор и соответствующего строения спороносного аппарата у мукоровых различают спорангиальный (эндогенный) и конидиальный (экзогенный) типы бесполого спороношения.
При спорангиальном типе (спорангиоспоры, мероспорангиоспоры) споры возникают в особых споровместилищах, образующихся на верхушке спороносцев (боковых веточек), которые бывают трех типов: спорангии, стилоспорангии и мероспорангии.
Спорангии - споровместилища более примитивные по своему строению. Это мелкие (до 80-120 мкм в диаметре) шаровидные вздутия, содержащие от одной до нескольких тысяч спор. При этом споры заполняют всю внутреннюю полость спорангия.
Стилоспорангии имеют чаще шаровидную форму. Они более крупные (достигают до 1,5 мм в диаметре), чем спорангии, и являются только многоспоровыми (содержат до 70 000 спор). Кроме того, стилоспорангии снабжены колонкой, которая представляет собой стерильную часть, ограниченную оболочкой, и которая является как бы продолжением верхушки спороносца (стилоспорангиеносца) в полости спорангия. Отсюда и само название - стилоспорангий (спорангий с колонкой). Колонка бывает шаровидной, цилиндрической, конической или обратногрушевидной, что имеет видовое систематическое значение.
Как в спорангиях, так и в стилоспорангиях образуются споры, именуемые спорангиоспорами. Они беспорядочно заполняют эти споровместилища и освобождаются из них в результате растворения или разрыва их оболочки.
Мероспорангий - это спорангий цилиндрической формы, расчленяющейся по поперечным перегородкам (перетяжкам) на "цепочку" спор (мероспорангиоспор). Мероспорангии, как правило, имеют мало спор и сохраняют оболочку, которая срастается с таковой боковых сторон мероспорангиоспор.
Половой процесс у мукоровых грибов зигогамный (гаметангиогамный), при котором происходит слияние содержимого (копуляция) двух особых клеток (гаметангиев) одного или разных талломов и образование так называемой зигоспоры. Основными элементами в этом способе размножения являются копулирующие отроги, выполняющие половую функцию, и зигоспора - продукт полового процесса.
Копулирующие отроги представляют собой обычно короткие боковые ответвления гиф. Они одноклеточные и морфологически отличаются от вегетативных гиф или не отличаются от них. Оба отрога могут быть одинаковые (виды изогамные) или неодинаковые (виды гетерогамные).
Зигоспоры - это одноклеточные образования, чаще шаровидной формы, с многослойной оболочкой. Наружный слой оболочки зигоспор обычно окрашенный и с бородавчатыми, пирамидальными или звездчатыми выступами. Из-за этих выступов кажется, что зигоспоры снабжены утолщениями соответствующей формы. У некоторых мукоровых грибов зигоспоры окружены сетчатым или войлочным покровом. В первом случае этот покров образован придатками копулирующих отрогов, а во втором - ответвлениями вегетативных гиф.
Зигоспоры образуются одиночно в месте соединения верхушек копулирующих отрогов. У отдельных мукоровых грибов зигоспоры развиваются на одном отроге. Такие партеногенетические зигоспоры принято называть азигоспорами.
Развитие полового спороношения начинается с возникновения у гиф боковых ответвлений - копулирующих отрогов, которые растут чаще навстречу друг другу, соединяясь своими верхушками. При этом в каждом отроге возникает поперечная перегородка, отделяющая небольшую верхушечную клетку - гаметангий, содержащий в цитоплазме небольшое число ядер. Затем оболочка у верхушки отрогов в месте их соединения растворяется, в результате чего содержимое гаметангиев сливается в образовавшейся при этом крупной клетке. Последняя увеличивается в размере и приобретает толстую оболочку в результате послойного отложения соответствующих веществ, превращаясь таким образом в зигоспору. Одновременно с формированием зигоспоры увеличиваются в размере и копулирующие отроги.
Слияние ядер в образовавшихся зигоспорах происходит вскоре после их формирования, перед прорастанием или в период их прорастания. Лишь немногие зигоспоры способны прорастать, и прорастают они обычно фруктификативно и только после некоторого периода покоя.
Моноспоровые (односпоровые) культуры большинства мукоральных грибов не развивают половое спороношение. Но если две или несколько таких культур одного вида выращивать совместно, тогда при определенных комбинациях скрещивания они образуют узкую зону зигоспор в месте встречи колоний. Такая особенность этих грибов связана с тем, что они существуют в двух отдельных половых формах (мужская и женская). Этот феномен открыт в микологии Блексли (1904) на примере мукоровых и известен как гетероталлизм. Виды, обладающие такой особенностью, именуются гетероталличными. Наряду с ними в порядке мукоровых есть виды, моноспоровые культуры которых постоянно образуют зигоспоры. Развитие последних при благоприятных условиях происходит у них более или менее равномерно по всей колонии. Такие мукоровые грибы, существующие в одной двуполой форме, принято называть гомоталличными.
У большого числа гетероталличных мукоровых грибов встречаются также такие особи, которые не развивают зигоспоры с половыми формами и не вступают в половую реакцию. Эти грибы нейтральны в половом отношении. Все эти формы гетероталличных мукоровых грибов возникают при митотическом делении ядер зигоспор, которые являются двуполыми. Такое расщепление вида происходит при прорастании зигоспор.
Как у гомо-, так и у гетероталличных видов пол обозначается знаком + или -. Причем у гетероталличных грибов под + и - понимается различие в поле отдельных культур, т. е. половые формы, а у гомоталличных этими знаками обозначаются только копулирующие отроги в пределах одной культуры, которая является обоеполой формой.
Мукоровые грибы - в основном сапрофиты, только небольшое число их ведет паразитический образ жизни, причем ограниченный, как правило, своими же сородичами.
Мукоровые грибы легко выделяются и хорошо развиваются в чистой культуре на агаровых средах, в особенности на органических, образуя чаще светлые пушистые колонии.
Мукоровые грибы широко распространены в природе. Они постоянно обитают в почве, где участвуют в процессах разложения органических веществ. В частности, они активно вызывают минерализацию белковых веществ растительного происхождения, при которой происходит обогащение почвы аммонийным азотом.
Другим классическим субстратом для мукоровых грибов служат экскременты травоядных животных, в особенности помет грызунов, а также навоз лошадей и рогатого скота (за исключением коров). Характерно, что на экскрементах мукоровые грибы проявляют выраженную сукцессию в развитии, что связано с их образом жизни (питания). Так, первоначально на экскрементах развиваются сапрофитные грибы, а после того, как их развитие достигает апогея, начинают развиваться факультативные, а затем и облигатно паразитные виды.
Мукоровые грибы довольно часто развиваются на различных кормах и на всевозможных пищевых продуктах, вызывая их порчу (плесневение), а также сопутствуют человеку в его повседневной деятельности. Многие из них вызывают также порчу урожая при его хранении или являются возбудителями микоза (мукоромикоза) человека, сельскохозяйственных животных и домашних птиц. Некоторые из них обладают высокой ферментативной активностью или продуцируют β-каротин, жир, различные органические кислоты, а поэтому находят практическое применение.
Мукоровые грибы - аэробы, но некоторые из них способны расти в отсутствие свободного кислорода и в таком случае образуют молочную кислоту. Характерно, что во всех остальных случаях эта кислота представляет собой типичный продукт анаэробного обмена веществ организмов и наличие кислорода воздуха тормозит эту реакцию - явление, известное как реакции Пастера.
Предполагается, что мукоровые грибы происходят от более низкоорганизованных грибрв (сапролегниевых, питиевых), развиваясь в сторону приспособления к наземным условиям существования.
В порядке мукоровых несколько семейств, различающихся преимущественно особенностями бесполого спороношения.
Семейство Мортиерелловые (Моrtierellaceae)
Это семейство объединяет грибы, имеющие только спорангии. Большинство его представителей, в отличие от остальных мукоровых грибов, имеют еще спороносцы (спорангиеносцы) шиловидной формы и неокрашенные колонии, нарастающие чаще лопастями, концентрическими зонами (сплошными или прерывистыми) или как радиально расходящиеся лучи (рис. 36, табл. 10).
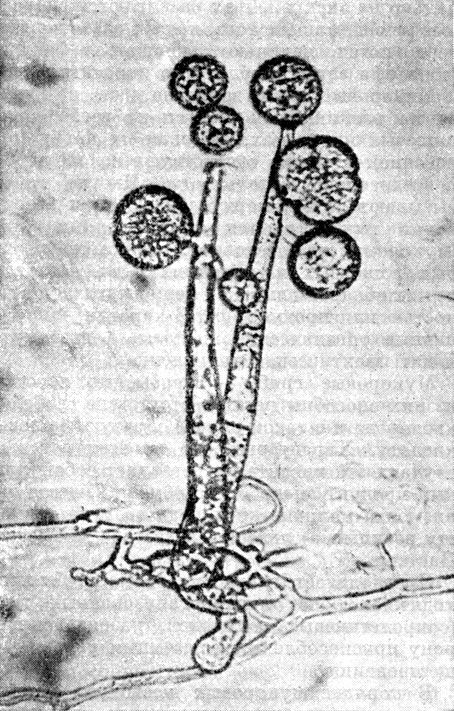
Рис. 36. Мортиерелла (Mortierella)
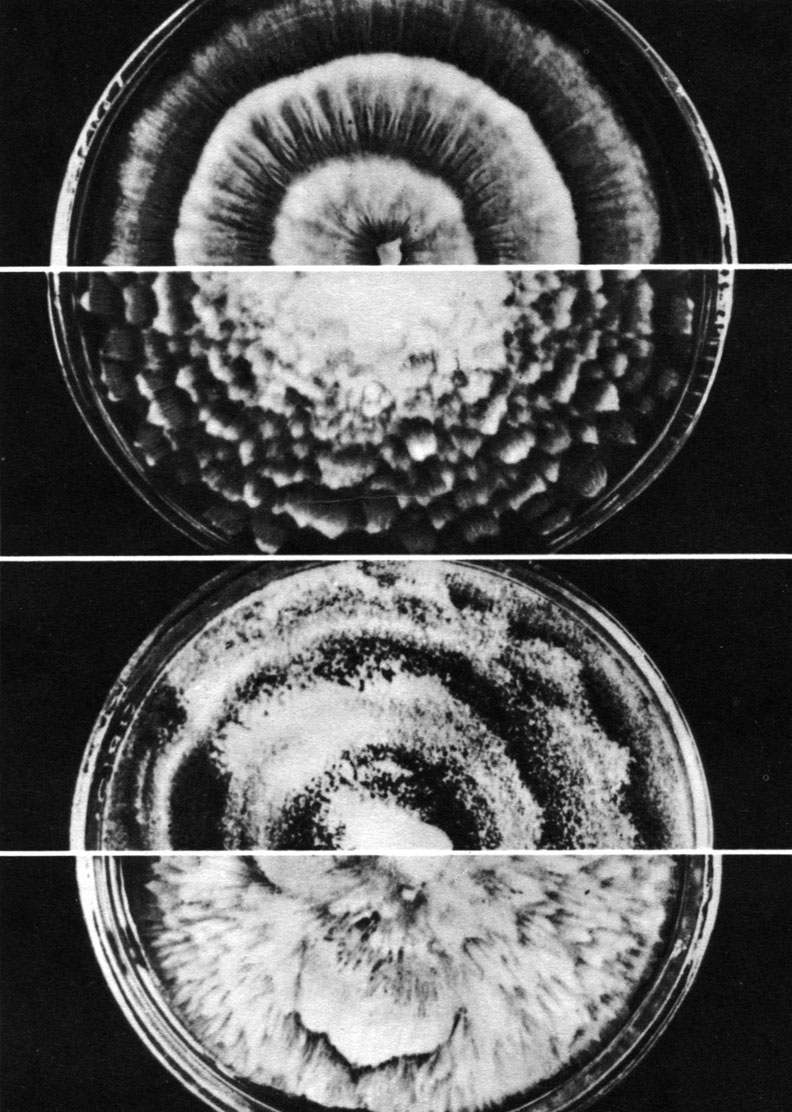
Таблица 10. Колонии разных видов рода мортиерелла (Mortierella) на агаре
Мортиерелловые грибы - сапрофиты, не имеющие народнохозяйственного значения. В семействе известно 3 рода, объединяющие до 70 видов.
Наиболее крупным в семействе является род мортиерелла (Mortierella), в котором около 65 видов. Грибы этого рода обитают чаще в торфе и во влажных почвах. В культуре они нередко издают чесночный запах, в особенности при выращивании их на среде с пептоном.
Семейство Пилоболовые (Pilobolaceae)
Семейство пилоболовых объединяет 3 рода с 15 видами. Пилоболовые грибы характеризуются простыми стилоспорангиеносцами и крупными отделяющимися стилоспорангиями. Копрофильные формы развиваются в природе, как правило, по замкнутому кругу: экскременты животных (чаще травоядных) - пастбищные растения - пищеварительный тракт животных - экскременты животных. Наиболее известны роды пилоболус, или пилобол (Pilobolus) и пилаира (Pilaira).
В роде пилоболус 8 видов. У грибов этого рода стилоспорангии имеют полушаровидную форму и резко отбрасываются по направлению к источнику света, отлетая вместе с колонкой. Характерно для этих грибов развитие стилоспор ангиеносцев из трофоцисты и вздутие их в верхней части (подспорангиальное вздутие). Предполагают, что это вздутие (табл. 11) выполняет функцию тонкого приспособления в наводке стилоспорангия по направлению к источнику света и в освобождении его.
Отбрасыванию стилоспорангиев у пилоболов предшествует повышение тургора в стилоспорангиеносцах. В результате повышения тургора наступает критический момент, при котором оболочка подспорангиального вздутия не выдерживает давления и мгновенно разрывается по окружности у основания колонки, а содержимое стилоспорангиеносца под давлением резко выбрасывается через образовавшееся отверстие тонкой струей. Стилоспорангиеносцы при этом мгновенно сморщиваются и спадают в результате внезапного падения тургора и потери содержимого. Начальная скорость полета стилоспорангиев примерно 4,7-27,5 м/сек, а летят они на расстояние до 2 м.

Таблица 11. Спорангий мукорового гриба пилобола (Pilobolus)
Отлетевшие стилоспорангии наталкиваются в полете на преграду и прикрепляются к ней слизью, освободившейся из этих стилоспорангиев после разрыва оболочки от удара. В природе стилоспорангии попадают на пастбищные растения. Затем их поедают животные, которые проглатывают и прикрепленные к растениям стилоспорангии со спорами. Эти споры проходят через пищеварительный тракт животных и выделяются с экскрементами. Затем на экскрементах споры прорастают и начинается новый цикл развития гриба.
В отличие от остальных мукоровых грибов пилоболы развиваются обычно только при щелочной реакции среды и для своего развития нуждаются в ростовом веществе - карпогене. Это вещество содержится в тканях травянистых растений и в экскрементах травоядных животных. Из пилоболов наиболее распространены пилоболус кристаллический (P. crystallinus), пилоболус длиннотрофоцистный (P. longipes) и пилоболус росянистый (P. roridus). Эти грибы встречаются чаще на конском навозе, где обильно развивают спороношение в утренние часы. При этом навоз как бы покрывается рыхлым войлоком, усеянным многочисленными мелкими бесцветными каплями.
Грибы рода пилаир а отличаются от представителей предыдущего рода тем, что у них нет трофоцисты, а стилоспорангиеносцы без подспорангиального вздутия и стилоспорангии вносятся на окружающую растительность в результате мгновенного вытягивания стилоспорангиеносцев. В СССР отмечены пилаира необычная (P. anomala) и пилаира кавказская (Р. caucasica).
Семейство Мукоровые (Mucoraceae)
Это семейство объединяет грибы, у которых стилоспорангии не отделяются. Освобождение спорангиоспор у мукоровых происходит в результате растворения или разрыва оболочки стилоспорангия непосредственно на стилоспорангиеносце. Стилоспорангии чаще шаровидные, и только у двух монотипных родов они колбовидные (род саксенея - Saksenaea, рис. 37) или сосисковидные (род эхиноспорангий - Echinosporangium, табл. 9).
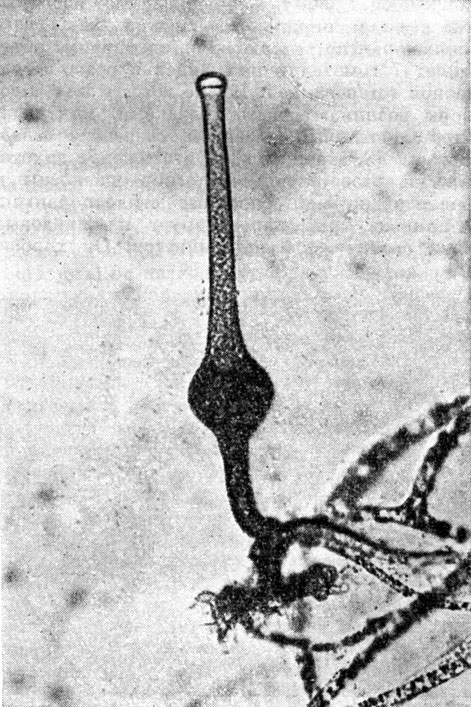
Рис. 37. Саксенея колбовидная (Saksenaea vasiformis)
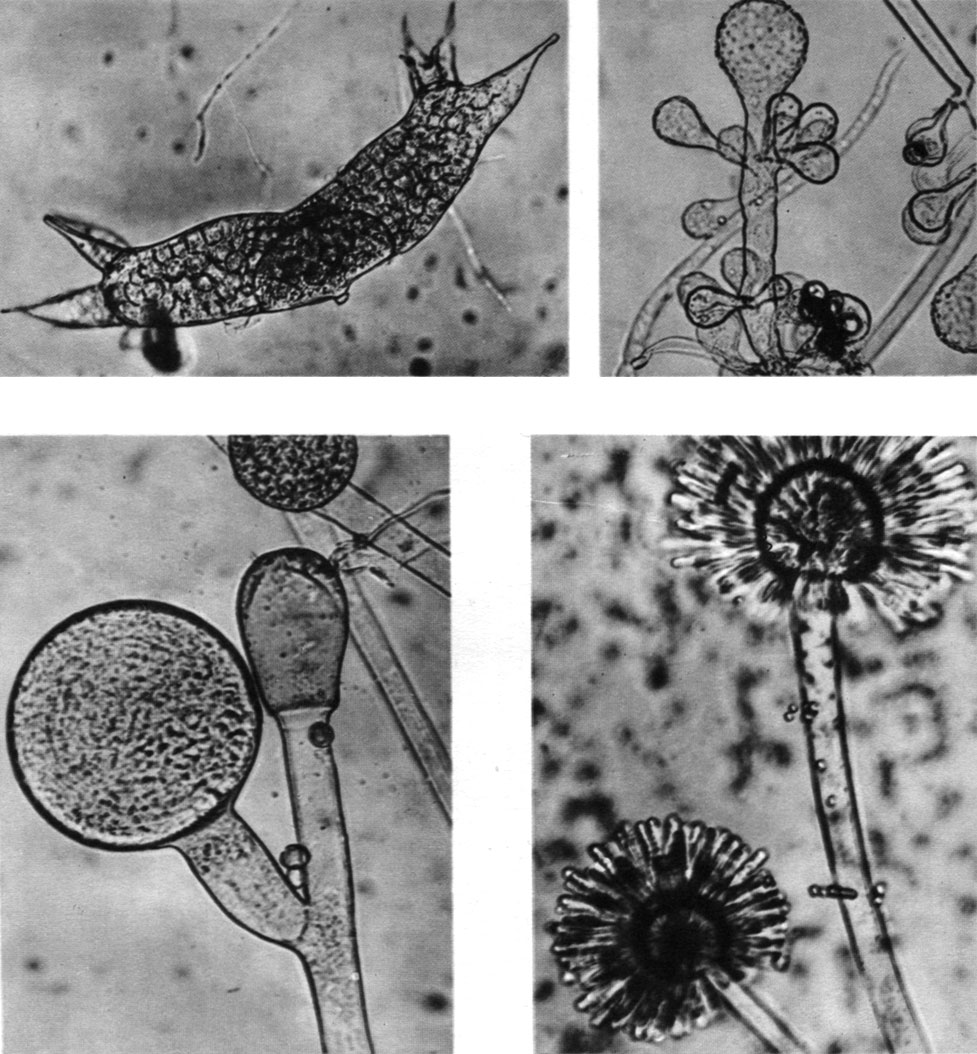
Таблица 9. Спороношения мукоровых грибов: вверху слева - эхиноспорангий поперечный (Echinosporangium transversalis); вверху справа - куннингамелла эхинулята (Cunninghamella echinulata); внизу слева - мукор петринсулярис (Mucor petnnsulans); внизу справа - синцефаляструм кистевидный (Syncephalastrum racemosum)
Мукоровые - самое крупное семейство в порядке. В нем 12 родов, охватывающие около 145 видов. Эти грибы широко распространены в почве, на экскрементах травоядных животных и на всевозможных гниющих субстратах растительного происхождения. Громадное большинство из них - сапрофиты.
Грибы некоторых видов семейства мукоровых обладают высокой ферментативной (главным образом амилолитической и протеолитической) активностью, благодаря чему находят практическое применение (в частности, в странах Азиатского континента). Таковы актиномукор изящный (Actinomucor elegans), ризопус малоспоровый (Rhizopus oligosporus), ризопус рисовый (R. oryzae), ризопус японский (R. japonicus), мукор китайский (Mucor sinensis), мукор кистевидный (М. racemosus) и мукор улитковидный (М. circinelloides).
Эти грибы используются в странах Азиатского континента в качестве компонента закваски ("китайские дрожжи", "раги") или непосредственно для ферментативного производства сброженных продуктов питания ("соевый сыр", "темпе" и т. п.) из бобов сои, зерна злаков (риса и др.), ядра кокосового ореха, а также для получения спирта из клубней картофеля. Некоторые мукоровые грибы вызывают микозы (мукоромикозы) легких ("ложный туберкулез"), головного мозга и других органов человека, сельскохозяйственных животных и домашних птиц, ассоциируются с гематурией и микотическим абортом у свиней, рогатого скота. Такой способностью, например, обладают абсидия ветвистая (Absidia ramosa), абсидия зонтикокистевидная (A. corymbifera), мукор мелкий (Mucorpusillus), ризопус Кона (Rhizopus cohnii) и ризопус рисовый (R. orysae). Эти грибы часто вызывают также порчу (плесневение, самосогревание, мокрую гниль) кормов (сена, соломы и др.), заскирдованных во влажном состоянии, а также гниение зерна и различных мясистых плодов (яблоки, груши, клубни картофеля, корнеплоды сахарной свеклы, ириса и др.) при хранении.
В СССР обнаружены представители большинства родов этого семейства.
Род мукор (Mucor) занимает центральное место в семействе. Грибы этого рода характеризуются одиночными бесцветными стилоспорангиеносцами. Они отходят от морфологически недифференцированных вегетативных гиф субстратного и (или) воздушного мицелия. Стилоспорангиеносцы могут быть простыми или различно разветвленными (чаще моноподиально, неправильно симподиально или кистевидно). У грибов с разветвленными стилоспорангиеносцами веточки обычно прямые, в связи с чем стилоспорангии прямостоячие. Большинство видов мукора гетероталличны.
В дополнение к указанным уже видам этого рода необходимо отметить еще М. раманниановый (М. ramannianus). Этот гриб очень широко распространен в кислых лесных почвах. В культуре гриб образует шерстистые колонии красновато-кирпичного цвета, и они часто с узкой концентрической зональностью. Отдельные культуры гриба образуют антибиотик рамицин.
Близки к роду мукор роды актиномукор (Асtinomucor), цирцинелла (Circinella), зигоринхус (Zygorhynchus) и сизигитес (Syzygites). Первый из них с видом актиномукор изящный (A. elegans) и характеризуется мутовчатым ветвлением стилоспорангиеносцев. Цирцинелла (в этом роде известно до 12 видов), в свою очередь, характеризуется симподиальным ветвлением спорангиеносцев. При этом боковые веточки спорангиеносцев обычно крючковидно изогнуты или в виде спирального завитка, из-за чего стилоспорангии свисают. Род зигоринхус объединяет 7 гомоталличных видов с резко выраженной гетерогамией. Кроме того, у них зигоспоры возникают часто гроздьями. Род сизигитес монотипный, с видом сизигитес аспергилловый (S. aspergillus). У этого гриба дихотомически разветвлены стилоспорангиеносцы и сравнительно многоспоровые стилоспорангии. В природе гриб паразитирует на плодовых телах различных базидиомицетов. Он хорошо развивается в культуре на органических агаровых средах, образуя пушисто-войлочные колонии серого цвета.
Род абсидия (Absidia) объединяет около 25 видов. Для этих грибов характерно наличие апофизы, из-за чего стилоспорангии имеют будто обратногрушевидную форму. Кроме того, у них обычно хорошо выражены столоны и стилоспорангиеносцы отходят чаще по нескольку от верхушки дуги столона (рис. 42).
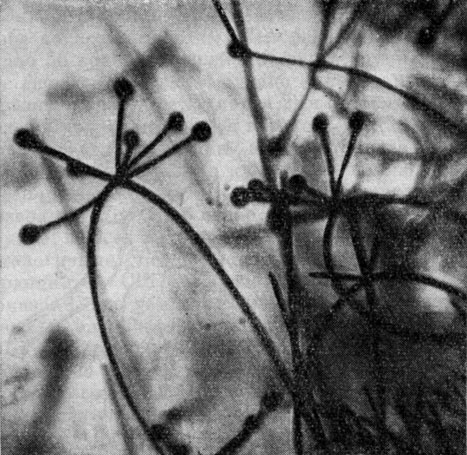
Рис. 42. Абсидия (Absidia)
Род ризопус (Rhizopus) содержит 11 видов. Они характеризуются темными стилоспорангиеносцами, которые отходят чаще по нескольку от шейки ризоида. У большинства видов хорошо развиты и столоны. В быту грибы этого рода известны под сборным названием "головчатая плесень" или "черная плесень". Многие представители его обладают высокой ферментативной активностью или образуют различные органические кислоты, благодаря чему находят практическое применение, в особенности в странах Азиатского континента.
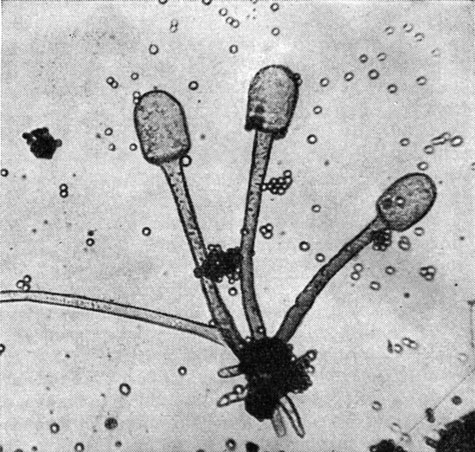
Рис. 41. Ризопус Кона (Rhizopus cohnii)
Близки к роду ризопус роды фикомицес (Phycomyces) и спинеллус (Spinellus), насчитывающие соответственно 3 и 6 видов. У грибов этих родов нет столонов, а стилоспорангиеносцы отходят одиночно от гиф субстратного мицелия. При этом представители рода фикомицес - безобидные сапрофиты, распространенные в зонах с теплым и умеренным климатом. Грибы рода спинеллус ведут паразитный образ жизни на плодовых телах различных базидиомицетов и распространены преимущественно в зоне с прохладным климатом. Кроме того, у фикомицеса, в отличие от спинеллуса, копулирующие отроги снабжены разветвленными шиповидными придатками, образующими сетчатый покров зигоспор. Фикомицес интересен тем, что его можно использовать как продуцента β-каротина.
Семейство Тамнидиевые (Thamnidiасеае)
Это небольшое семейство объединяет грибы, у которых наряду со стилоспорангиями развиваются также конидии и (или только) спорангии. Причем спорангии и (или) конидии образуются на отдельных спороносцах (спорангиеносцах, конидиеносцах) или совместно со стилоспорангиями на стилоспорангиеносцах. В семействе 6 родов, из которых у монотипных радиомицес (Radiomyces) и кокеромицес (Соkeromyces) стилоспорангии отсутствуют. Бесполое спороношение у них представлено только спорангиями. Наиболее известны (в том числе и в СССР) роды тамнидий, хоанефора и геликостилум.
Род тамнидиум (Thamnidium) объединяет 4 вида, характеризующиеся дихотомически разветвленными спорангиеносцами с верхушечными спорангиями и обычно простыми стилоспорангиеносцами. Близким к нему является род геликостилум (Helicostylum) с 8 видами. Для представителей этого рода характерны боковые спорангии и стерильные придатки на стилоспорангиеносцах, которые обычно хорошо разветвлены (рис. 38). Грибы обоих родов встречаются чаще в почве и на экскрементах травоядных животных. Отдельные их представители хорошо развиваются при низких температурах и вызывают плесневение говяжьих мясных продуктов при хранении в холодильных установках. Такой особенностью характеризуются, например, тамнидиум изящный (Т. elegans) и геликостилум прекрасный (Н. pulchrum).
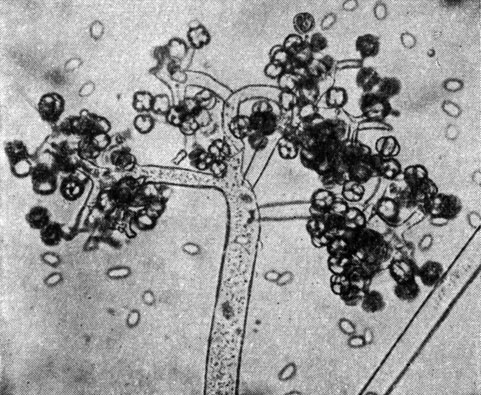
Рис. 38. Тамнидиум изящный (Tamnidium elegans)
В роде хоанефора (Choanephora) 10 видов, у которых спорангиоспоры с пучком щетинок на двух противоположных концах. Некоторые из грибов рода слабо и спорадически развивают стилоспорангии, но зато обильно образуют спорангии или конидии. Последние, как правило, табачного цвета и продольно исчерчены. Представители этого рода распространены в зонах с жарким и теплым климатом, где развиваются обычно в садовой и огородной почве. Большинство из них поражает цветки и плоды различных огородных и косточковых плодовых культур, причем поражаются ими эти органы у ослабленных растений или при наличии у них механических повреждений. Отдельные виды рода продуцируют β-каротин, выход которого резко повышается в случае совместного выращивания "мужской" и "женской" половых форм. Наиболее активным продуцентом каротина является хоанефора триспоровая (С. trispora).
Семейство Пиптоцефалидиевые (Piptocephalidaceae)
В это семейство входят грибы с мероспорангиальным типом бесполого спороношения. Мероспорангии возникают на плодущих вздутиях мероспорангиеносцев (боковых веточках) или на их особых плодущих структурах (спорокладиях) по нескольку, образуя шаровидные головки или короткие колонки. В семействе 3 рода.
Род синцефаляструм (Syncephalastrum) монотипен, в нем один вид - синцефаляструм кистевидный (S. racemosum). Гриб синцефалястр ведет сапрофитный образ жизни и характеризуется симподиально разветвленными мероспорангиеносцами (табл. 9).
Грибы родов синцефалис (Syncephalis) и пиптоцефалис (Piptocephalis) являются облигатными паразитами, развивающимися, как правило, на других мукоровых грибах, в особенности на копрофильных формах. Причем род синцефалис (27 видов) характеризуется простыми мероспорангиеносцами и мероспорангиоспорами, погруженными в слизь, в то время как у рода пиптоцефалис (17 видов) мероспорангиеносцы, многократно дихотомически разветвленные, и мероспорангиоспоры, как правило, не погружены в слизь (виды сухоспоровые). Кроме того, грибы синцефалиса, в отличие от грибов рода пиптоцефалис, сильно угнетают развитие гриба-хозяина (рис. 39, 40).
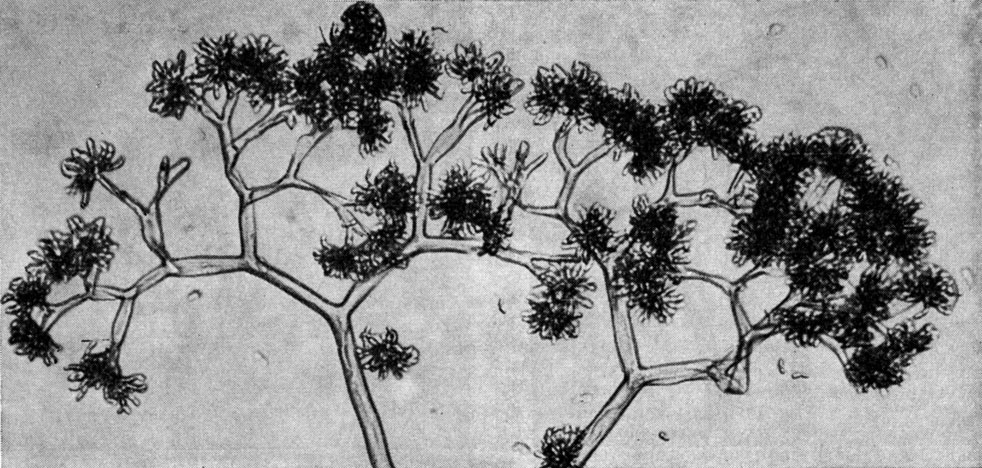
Рис. 39. Пиптоцефалис (Piptocephalis)
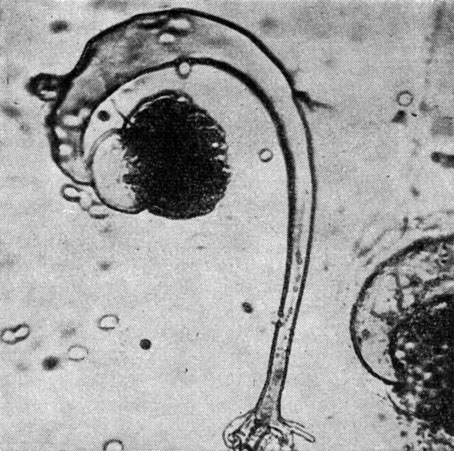
Рис. 40 Синцефалис роговой (Syncephalis corni)
Семейства Куннингамелловые (Cunninghamellaceae), Кикселловые (Kickxellaceae) и Димаргаритовые (Dimargaritaceae)
Грибы этого семейства характеризуются конидиальным типом бесполого спороношения. При этом конидии возникают одиночно на плодущих вздутиях конидиеносцев (боковых веточках), образуя, в зависимости от формы вздутий, шаровидную головку или колонку. В семействе известно 6 родов, объединяющих 16 видов. Это сапрофитные формы, распространенные довольно широко в почве и на отмерших субстратах растительного происхождения.
Наиболее известны роды куннингамелла (Cunninghamella) и хетоклядиум (Chaetocladium). Отдельные представители хетоклядиума, подобно грибам геликостила, вызывают плесневение говяжьих мясных продуктов при хранении в холодильниках.
Грибы остальных двух семейств порядка мукоровых - кикселловые (Kickxellaceae) и димаргаритовые (Dimargaritaceae) - ведут обычно паразитный образ жизни (чаще на других мукоральных) и известны преимущественно на Американском континенте.
Семейство кикселловые (в нем известно 3 рода с 12 видами) объединяет формы, характеризующиеся одиночными конидиями. Последние, в свою очередь, образуются более или менее тесным слоем на спорокладиях, которые имеют чаще лодковидную форму.
Семейство димаргаритовые, в свою очередь, содержит 7 родов, насчитывающих 22 вида. У грибов этого семейства конидии собраны по две в акропетальной цепочке и возникают мутовками на верхней части клеток спороклядий. Спороклядии у них состоят из 2-4 клеток, собранных в цепочки.
|
ПОИСК:
|
При использовании материалов проекта активная ссылка обязательна:
http://gribochek.su/ 'Библиотека о грибах'