Урожаи грибов и их цикличность
Под урожайностью макромицетов в микологии понимают массу видимой их части - спорадически появляющихся плодовых тел грибов, но ведь основная, рабочая часть гриба - мицелий - скрытая от наших глаз в субстрате: лесной подстилке, опаде, древесине, почве. Грибные гифы пронизывают мертвое органическое вещество до уровня внутриклеточных структур, в связи с чем до сих пор не удается достоверно определить запасы мицелия макромицетов ни в лабораторных условиях, ни тем более в природе. Это касается не только грибов, но и многочисленных обитателей лесной подстилки и почвы: водорослей, беспозвоночных животных, микромицетов, бактерий, простейших. Что касается макромицетов, имеются немногочисленные литературные сведения о соотношении массы мицелия и массы плодовых тел, рассчитанные на основе экспериментов, проведенных в лабораторных условиях. Для напочвенных сапротрофов это соотношение составляет 62,6:1, а для симбиотрофов 154:1, т. е. для формирования одного грамма плодового тела гриба в первом случае необходимо 62,6 г мицелия, во втором - 154. Эти цифры внушительны, но сколько же необходимо переработать субстрата, чтобы создать один грамм мицелия? Это пока неизвестно, как и неизвестно физиологическое состояние мицелия в природе, перераспределение вещества и энергии между плодовым телом гриба и мицелием. Исходя из приведенных цифр, мы для ельника Подмосковья (возраст 80 лет, примесь в древостое березы и дуба) рассчитали массу мицелия макромицетов. Она оказалась равной 2,5 т на 1 га и превосходит по величине массу мхов и травянистых растений. Доля мицелия грибов и других подстилочнообитающих организмов по массе составляет 25% от запасов опада и подстилки в этом лесу. Эта цифра может быть и выше, а это значит, что в лесу мы буквально ходим "по живому", не подозревая, какой удивительный мир организмов у нас под ногами скрыт от наших глаз опавшими листьями, травой, мхами!
Для конкретного леса основную массу плодовых тел формируют симбиотрофы и ксилотрофы, обладающие крупными плодовыми телами, хотя по количеству видов они могут и уступать напочвенным сапротрофам. Колебания урожаев в одном типе леса могут достигать огромных величин в многолетней динамике в основном за счет периодичности массового плодоношения микоризообразователей и дереворазрушающих грибов. Причины периодичности урожаев съедобных грибов обычно связывают с количеством осадков и температурой конкретного года. Имеются даже попытки прогнозировать урожайность грибов, но поскольку прогноз погоды у нас еще не совсем объективен, предсказание урожаев грибов не состоятельно.
При исследовании зависимости урожаев грибов от гидротермических условий учитывают количество осадков, температуру воздуха и почв, предшествующих или совпадающих по времени с их интенсивностью. Однако при этом совершенно не учитываются физиологические циклы развития мицелия, которые наверняка играют ничуть не меньшую роль, чем погодные условия. По нашим наблюдениям, малосходные погодные условия вызывали массовое появление в лесах Подмосковья веселок, ежовика коралловидного с периодичностью соответственно 8 и 5 лет. Речь идет именно о массовом их появлении, так как отдельные экземпляры этих видов изредка встречались и в годы между циклами. Игнорирование физиологической цикличности плодоношения грибов вносит бесконечные разногласия в вопросе о ведущих факторах, определяющих урожайность грибов, а зачастую приводит и к необоснованным суждениям о редкости видов.
Физиологическая цикличность урожаев грибов в чистом виде должна проявляться только в годы, приближающиеся по количеству тепла и влаги к среднемноголетнему, в достаточной степени абстрактному вегетационному периоду. Существенную поправку в этот ритм, безусловно, вносят погодные условия конкретного года, давая толчок к массовому плодоношению либо более сухолюбивых, либо, наоборот, влаголюбивых видов.
Чтобы разобраться в вопросе, что же определяет массовое появление грибов - погодные условия или физиологическая цикличность, или сочетание обоих фактов, рассмотрим ритмичность плодоношения опенка настоящего яли осеннего - одного из самых распространенных и уважаемых грибниками представителей макромицетов. Наблюдения проводились в лесах Подмосковья, но прежде чем переходить к исследованию закономерностей массового появления опенка, необходимо остановиться на некоторых биологических особенностях самого гриба. Осенний опенок космополитный вид, произрастающий на древесине более 200 видов растений, в основном деревьев и кустарников. На территории СССР он встречается в различных природных зонах на видах хвойных и лиственных пород - ели, сосны, пихты, лиственницы, березы, осины, дуба, липы, клена, ясеня, каштана, яблони, сливы, сирени, а также на кустарниках - малине, крыжовнике, смородине. В качестве источника пищи опенок использует как представитель факультативных (необязательных) паразитов углеводы растительных тканей, в большинстве случаев камбия. Оптимальные условия для распространения мицелия и ризоморфов (собранные в темноокрашенные шнуры гифы гриба различной толщины и до нескольких метров длины), а также обилие плодовых тел опенка связывают с влажностью почв и концентрацией кислорода в них. Чем выше содержание кислорода и соответственно ниже содержание углекислого газа, тем выше границы распространения ризоморфов в почве. Во влажных местообитаниях основная масса ризоморфов сосредоточена в верхнем 10-сантиметровом слое почвы. Развитие мицелия в субстрате зависит также от содержания и соотношения в нем углерода и азота. Причем повышенное содержание азота способствует развитию ризоморфов и стимулирует паразитарную активность опенка, а возрастание количества углерода вызывает активное разрастание воздушного мицелия, определяя сапротрофный образ жизни гриба.
В природных условиях опенок может быть и микоризообразователем (у орхидных), и сапротрофом, разлагающим только мертвую древесину, и опасным паразитом, уничтожающим целые массивы лесов. Вопрос о принадлежности опенка осеннего к группе сапротрофных или паразитных дереворазрушающих грибов дискутируется издавна, но большинство советских микологов и лесоводов относят его к факультативным паразитам. Скорее всего, это так, потому что паразитизм опенка проявляется только в определенных, неблагоприятных для деревьев и кустарников условиях, когда они находятся в ослабленном жизненном состояния (высокий уровень корневой конкуренции между деревьями за питательные элементы, влагу, свет, различного рода повреждения корней и стволов).
Мы изучали распределение и урожайность опенка осеннего в смешанных (елово-лиственных) лесах, осинниках и березняках, искусственных насаждениях ели и сосны различного возраста, вырубках Подмосковья в течение нескольких лет. Опенок в эти годы встречался во всех лесах выше 30-летнего возраста и на вырубках различного возраста. Максимальное количество плодовых тел гриба отмечено во все годы на пнях и валежной древесине ели и березы, на сухостойных деревьях ели, березы, дуба, ивы, лещины (лесной орех). Изредка в отдельные, особенно благоприятные и урожайные, годы отмечалось нахождение опенка на пнях и сухостое осины, черемухи, сосны, сирени. На древесине липы и клена плодовых тел гриба не найдено.
В качестве паразита опенок в названных лесах встречался в основном на корнях и у основания стволов ели в искусственных ее посадках в возрасте от 40 до 80 лет. Большое количество плодовых тел сплошь покрывает нижние части стволов деревьев, от уровня земли до высоты одного метра. Это темные леса с почти полным отсутствием напочвенного травяного покрова. Стоящие в рядах деревья (расстояние между рядами около 5 м, между деревьями в рядах - 3 м) к указанному возрасту оказываются в условиях жесткой конкуренции, следствием чего является угнетенное, ослабленное состояние отдельных деревьев. В тканях таких деревьев происходят и физиологические изменения: снижается активность окислительно-восстановительных процессов, уменьшается количество аминокислот и глюкозидов в корнях. На корнях и у основания деревьев, находящихся в более благоприятных условиях (опушки посадок, зоны их контакта с просеками, лесными дорогами), опят не обнаружено. Механическое же нарушение корневой системы деревьев (обрубка корней) моментально (на первый, второй год) вызывает массовое появление опенка в зоне повреждения. Такая быстрая реакция подтверждена экспериментами с выращиванием опенка в культуре (лабораторные условия), когда на мицелии, выращенном из спор, плодовые тела образовывались через 2-5 мес на древесине ели и сосны.
Реже опенок встречается на живых деревьях березы и дуба, и то только тогда, когда они находятся на гранях своего жизненного уровня (возраста) или механически повреждены, или повреждены жуками-короедами. В качестве субстрата в этих случаях гриб использует отмершие части коры. В отдельные годы массового плодоношения опенок встречается на живом подросте ели, березы, дуба, где молодые деревца образуют непроходимые заросли.
Таким образом, опенок как факультативный паразит выступает в роли индикатора угнетенного состояния деревьев и способствует выпадению их из древостоя (изреживанию), выполняя роль санитара и биологического регулятора видового состава и численности растений. При выпадении отдельных деревьев образуются окна в древесном пологе, изменяются экологические условия для рядом стоящих деревьев, т. е. увеличивается площадь питания корневой системы, большее количество осадков проникает к поверхности почвы, изменяется освещенность, перестраивается напочвенный покров. Проникновение ультрафиолетовой части солнечной радиации к поверхности корневых лап и оснований стволов деревьев может служить причиной подавления паразитарной активности опенка.
Более типичным в названных лесах является сапротрофный тип развития опенка. Он встречается на пнях и валеже под пологом леса и на вырубках. Пни лиственных пород деревьев заселяются грибом обычно в первый же год их существования независимо от того, было дерево поражено паразитной формой опенка или не было. После рубки ели, в случае, если при жизни на ней не было опенка, появление плодовых тел гриба наблюдается не ранее, чем через 2 года и продолжается на протяжении 8 лет. На вырубках и просеках опенок плодоносит в течение 10-15 лет до почти полного разложения древесины, скорость которого зависит от экологической обстановки конкретного местообитания.
Так, на протяжении 1969-1980 гг. наблюдалось наряду с редким и единичным появлением опенка в определенных местообитаниях и на определенном субстрате массовое его плодоношение. Сроки начала плодоношения оценка осеннего в лесах Подмосковья колеблются в интервале 10 дней, с 26 августа по 7 сентября. Исключение составлял только 1972 год, необычайно жаркий и сухой, когда горели торфяники и леса в Московской, Рязанской, Владимирской, Горьковской, Костромской и других областях. В то лето грибов практически не было, температура подстилки в лесах в июле поднималась до невероятной отметки +37° и при полном отсутствии осадков мицелий многих грибов сильно пострадал, а некоторых погиб (напочвенных сапротрофов, обладающих мелкими плодовыми телами). Опенок в этот год появился лишь 18 сентября, после выпадения дождей в очень небольшом количестве. Многие микологи связывают годы массового плодоношения опенка с резкими перепадами температуры воздуха в конце лета и увеличением количества осадков, вызывающих высокие показатели относительной влажности воздуха. Исходя из этого, мы проанализировали погодичное количество осадков в течение вегетационного периода, т. е. изменение влажности субстрата до средних сроков появления плодовых тел опенка (к 1 сентября), количество выпавших осадков подекадно в августе, а также сумму положительных температур, накопленных к 1 сентября, изменения средних, минимальных и максимальных температур воздуха ежемесячно и подекадно.
Рассмотрим вначале зависимость урожайности опенка осеннего (Armillariella mellea) от погодичных изменений температуры. На рис. 8 показана величина, отражающая отношение результирующей накопленных положительных температур четырех месяцев (май - август) к сумме температур за эти месяцы. Она указывает на равномерность накопления тепла; чем ближе к 0°С, тем равномернее распределены температуры в течение исследуемого периода. Абсолютная величина результирующей колеблется от 25 до 320° соответственно в 1979 и 1972 гг. Максимальное количество накопленных положительных температур в июне наблюдалось только в 1969 г., в июле - в 1973, 1974, 1975, 1977 и 1979 гг., в августе - в 1970, 1971, 1972, 1976, 1978 и 1980 гг. Как видно из рисунка, накопление температур происходит с выраженными колебаниями минимума и максимума, период колебаний составляет 2 года. На этом же рисунке отмечены годы с массовым плодоношением опенка. Анализ показывает, что периоды колебаний исследуемых явлений не совпадают. Были рассмотрены и показатели накопленных к 1 сентября (началу плодоношения опенка) положительных температур. Во все исследуемые годы она к этому времени равнялась 2000-2500°. Различия эти определяются суммой температур март - конец июня. Месячные и подекадные средние, максимальные и минимальные температуры воздуха вариабельны, а их анализ не дает никаких оснований связывать их величину и изменчивость с периодичностью плодоношения опенка. Можно лишь отметить, что массовому появлению плодовых тел гриба соответствуют величины абсолютного минимума температур - от -1 до +3°, абсолютного максимума - от +18 до +22° в августе.
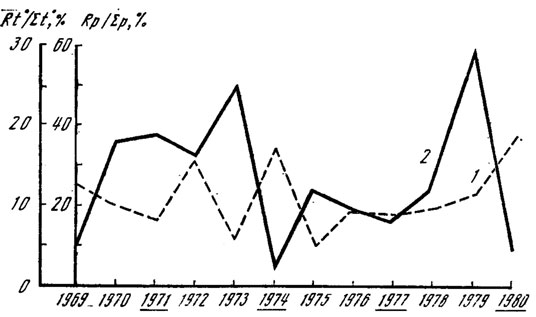
Рис. 8. Количество осадков, суммы положительных температур и плодоношение Armillariella mellea: Rt0 - результирующая температура месяцев (май - август); Rp - результирующая осадков тех же месяцев, мм; ∑р - сумма осадков, выпавших к 1 сентября; ∑t0 - сумма положительных температур, накопленных к 1 сентября; 1 - Rt0/∑t0, %; 2 - Rp/∑p, %, Подчеркнуты годы массового плодоношения опенка
Как было отмечено, наиболее резкие различия в накоплении суммы положительных температур наблюдаются в весенние месяцы и особенно в апреле. Амплитуда колебаний этой величины по годам достигает 275°, однако высоким урожаям опенка сопутствуют значения суммы положительных температур в очень узких пределах - 80-90°. Это единственная закономерность, которую нам удалось обнаружить.
Количество выпавших осадков в исследуемые годы резко различается в своем распределении по месяцам, декадам и сумме к 1 сентябри. Такого четкого и ритмичного колебания минимума и максимума, которое было отмечено для суммы положительных температур, в случае с осадками не наблюдается. Об этом свидетельствуют данные, представленные на рис. 9. Отношение результирующей четырех месяцев (май - август) к сумме осадков, выпавших за эти месяцы, указывает на резко выраженную их неравномерность. Причем никакой зависимости между массовым плодоношением опенка и распределением осадков не наблюдается. Однако можно отметить, что урожайным годам соответствует более или менее равномерное распределение осадков, когда величина отклонения их в различные месяцы не превышает 5-8%. Между абсолютным количеством осадков, выпавших к началу плодоношения опенка, а также ежемесячным и подекадным их режимами и появлением плодовых тел гриба зависимости не найдено. Некоторую информацию о связи этих явлений дает количество осадков, выпавших в последнюю декаду августа, перед непосредственным началом плодоношения. Сумма их колеблется от 3 до 5 в 1971, 1974 и 1977 гг. до 32 мм в 1980 г.
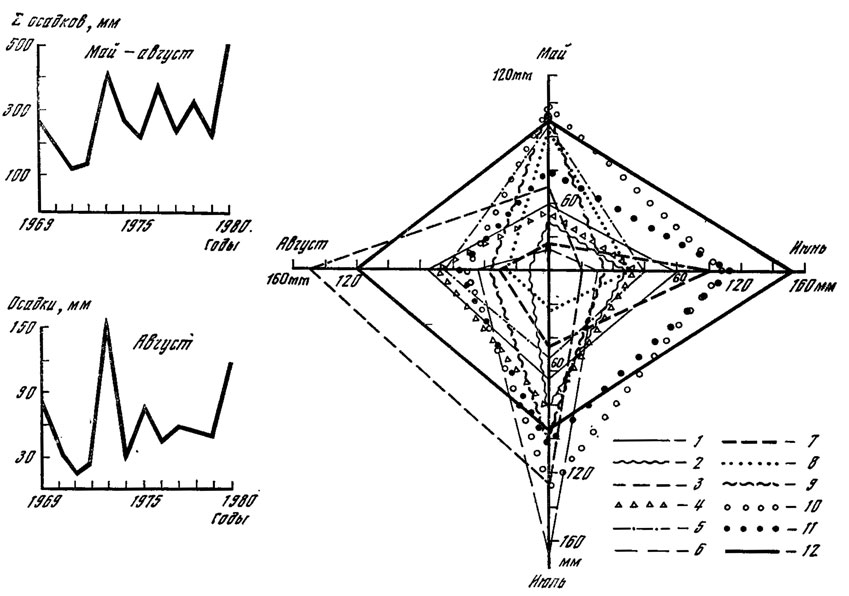
Рис. 9. Амплитуда колебаний выпавших в 1969-1980 гг. осадков: 1 - 1969; 2 - 1970; 3 - 1971; 4 - 1972; 5 - 1973; 6 - 1974; 7 - 1975; 8 - 1976; 9 - 1977; 10 - 1978; 11 - 1979; 12 - 1980
Одновременный анализ распределения температур и осадков в связи с цикличностью плодоношения опенка показывает, что урожайные годы характеризуются довольно резкими различиями в показателях суммы накопленных положительных температур и количества выпавших к 1 сентября осадков. В 1971 г. максимальное количество осадков выпало в июле на фоне равномерного накопления температур; 1974 г. (год массового появления опенка) характеризуется жарким июнем при влажном июле; 1977 г. - максимально влажным июнем и жарким июлем и, наконец, 1980 г. - самыми высокими показателями температуры и осадков в августе. Периодичность массового развития опенка, как видно из рис. 9, составляет 3 года и не совпадает с рассмотренными показателями температурного режима и режима влажности. По-видимому, такая цикличность урожаев опенка связана с иными причинами, такими, как, например, физиологические особенности развития гриба, в частности накопление достаточного количества веществ и энергии в мицелии.
Периодичность плодоношения макромицетов зависит от сочетания очень многих факторов. На рис. 10 показана многолетняя динамика количества видов и массы грибов различных экологических групп в одном и том же типе леса. Амплитуда колебаний массы грибов всех групп равна 14,8 кг/га, т. е. приближается к среднемноголетним значениям урожая плодовых тел. Такие различия определяются экстремальными годами, каковым оказался 1972 год. В основном же для лесов Подмосковья различия в урожайности грибов составляют 5-6 кг/га и совпадают по цикличности с суммой накопленных положительных температур за период май - сентябрь. Величина этого цикла - 2 года, а поскольку основу массы грибов составляют макромицеты с крупными плодовыми телами, относящиеся к симбиотрофам, можно считать, что периодичность урожаев таких грибов, как сыроежки, грузди, рядовки, паутинники, белые и березовики, составляют именно 2 года.
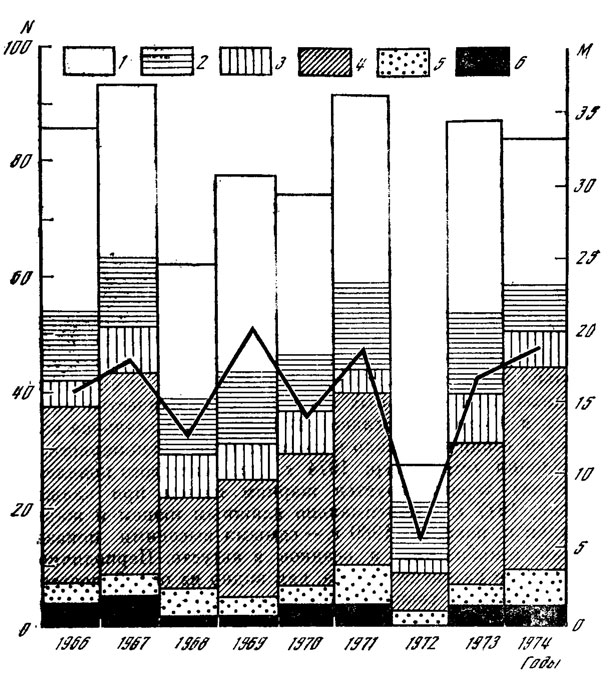
Рис. 10. Динамика трофической структуры и биомассы макромицетов (липо-ельник зеленомошно-волосистоосоковый). Число видов: 1 - симбиотрофов; 2 - ксилотрофов; 3 - гумусовых сапротрофов; 4 - подстилочных сапротрофов; 5 - сапротрофов на опаде; 6 - грибов других трофических групп. Жирной линией обозначена масса макромицетов, кг/га (абс. сухой вес); N - число видов; М - масса грибов
Наибольшей стабильностью в отношении видового состава, численности и массы обладают ксилотрофы и гумусовые сапротрофы, мицелий которых не испытывает в силу своего пространственного положения (защищенности) таких резких колебаний температуры и влажности, как мицелий грибов, расположенных в верхних слоях подстилки. Кроме того, как указывалось ранее, ксилотрофы имеют специфические приспособления для перенесения неблагоприятных погодных условий. Однако самыми консервативными в отношении количества видов и массы оказываются в многолетней динамике сапротрофы на опаде. Объясняется это уникальными физиологическими особенностями плодовых тел этих грибов. На опавших хвоинках и веточках хвойных пород постоянно присутствует в "увядшем" состоянии большое количество хрящеватых тонких (до одного миллиметра) ножек плодовых тел с зачатками шляпок (рис. 10). При повышении относительной влажности приземного слоя воздуха до 70-80% они тут же восстанавливают тургор тканей с минимальной затратой на этот процесс веществ и энергии.
Подстилочные сапротрофы наиболее уязвимы в отношении погодных условий, так как их мицелий, находящийся в различных слоях подстилки, слабо защищен от перепадов температуры и влажности, у них нет симбионтов, которые могли бы помочь им в защите, и нет специальных приспособлений, как у ксилотрофов (ризоморфов), для перенесения неблагоприятных условий.
Масса и количество видов макромицетов в течение одного сезона постепенно увеличиваются с мая, достигая максимальных значений в августе - начале сентября, а затем резко или медленно в зависимости от погодных условий конкретного года уменьшаются. Заканчивается плодоношение всех представителей экологических групп грибов в октябре - начале ноября. Самыми холодостойкими являются некоторые виды ксилотрофов и симбиотрофов, замерзающих на корню (опята, белые, рядовки).
Такое распределение массы макромицетов характерно для лесов умеренного климата. В районах с резко выраженным недостатком тепла за вегетационный период (на севере) и влаги (на юге) сроки роста, увеличения размеров и массы плодовых тел грибов укорачиваются по сравнению, например, с Подмосковьем в 2-3 раза. Реакция мицелия грибов на дефицитные тепло и влагу моментальная, почти как у сапротрофов на опаде. Здесь не каждый год создаются благоприятные погодные условия для образования плодовых тел грибов, но, когда это происходит, масса их огромна - 80-100 кг/га, а количество грибов на одном квадратном метре достигает 10 экз.
Обычно каждый гриб появляется в определенное для него время, однако в некоторые годы чисто погодные условия могут сдвигать сроки плодоношения. Так, например, в лесах Подмосковья понижение дневной температуры в июле 1976 г. до +10-12°, сопровождавшееся затяжными моросящими дождями, спровоцировало появление плодовых тел типично осенних видов - волнушки розовой и черного груздя. Отклонения в температуре и количестве осадков летом 1983 г. изменили все сроки развития грибов. Жаркая и влажная погода в июне - июле вызвала массовое развитие "колосовиков" - белых и березовиков и затянула их плодоношение на 1,5-2 мес. в отличие от среднемноголетнего (1,5-2 нед). Наступившая за этим жаркая и сухая погода августа исключила плодоношение всех симбиотрофов, связанных по срокам с этим месяцем.
Таким образом, на цикличность, величину урожаев, количество видов грибов влияет много факторов, среди которых основными являются физиологическое состояние мицелия и сочетание температуры и влаги в субстрате. Но ни в коем случае нельзя связывать урожайность макромицетов только с одним из них, например с осадками или температурой, как это зачастую у нас делается даже в научной (микологической) литературе.
|
ПОИСК:
|
При использовании материалов проекта активная ссылка обязательна:
http://gribochek.su/ 'Библиотека о грибах'