Порядок Микроасковые (Microascales) (И. И. Сидорова)
К небольшому порядку микроасковых относят грибы с темноокрашенными перитециями, содержащими беспорядочно расположенные прототуникатные сумки. Перитеции образуются на поверхности субстрата или частично в него погружены. Они мелкие, шаровидные или грушевидные, часто с длинным хоботком, в несколько раз превышающим по длине диаметр перитеция (Ceratocystis). В устье перитециев расположены перифизы, а парафизы отсутствуют. Оболочки сумок быстро лизируются, и зрелые перитеции содержат массу аскоспор, погруженных в слизь. При ее набухании аскоспоры вместе со слизью выходят из перитеция в виде слизистых капелек (Ceratocystis) или длинных слизистых шнуров (Microascus, рис. 85). Аскоспоры всегда одноклеточные, бесцветные или окрашенные.
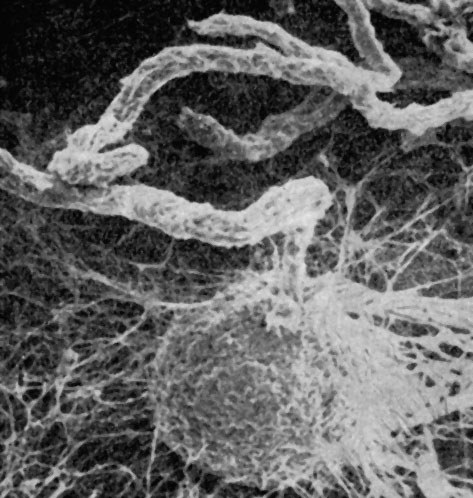
Рис. 85. Микроаскус (Microascus). Перитеций с выходящим из него шнуром аскоспор
Как и у эуроциевых, у микроасковых хорошо прослеживается морфологическая редукция полового процесса - переход от типичной гаметангиогамии до автогамии. У некоторых видов микроасковых, например у букового цератоцистиса (Ceratocystis fagacearum), впервые у аскомицетов наблюдается сперматизация. Сумки развиваются на аскогенных гифах по способу крючка (Microascus и Ceratocystis ulmi) или из голых, лишенных клеточных стенок аскогенных клеток (С. fagacearum).
У многих микроасковых в цикле развития преобладает конидиальное спороношение.
Микроасковые - сапрофиты на разнообразных растительных субстратах или паразиты высших растений, преимущественно деревьев. Некоторые из них вызывают серьезные болезни растений, приносящие большой ущерб сельскому и лесному хозяйству (голландская болезнь вязов, вилт дуба, гнили различных растений). Ряд видов микроасковых вызывает синеву древесины.
Род цератоцистис (Ceratocystis) из семейства офиостомовых (Ophiostomataceae) объединяет более 40 видов, распространенных как в умеренной зоне, так и в тропиках. Для грибов этого рода характерны перитеции с длинным хоботком, расположенные одиночно или группами на поверхности субстрата или погруженные в него основаниями. Их окраска варьирует от светло-коричневой до черной, хоботок обычно более темный. Диаметр перитециев 80-600 мкм, а длина хоботка - 150-9000 мкм.
Большое число видов этого рода паразитирует на растениях, чаще на деревьях (С. ulmi, С. fagacearum), лишь немногие виды встречаются на травянистых растениях.
К этому роду относятся такие экономически важные виды, как цератоцистис вязовой (С. ulmi) - возбудитель голландской болезни вяза; буковый (С. fagacearum), вызывающий вилт (увядание) дуба; бахромчатый (С. fimbriata) - возбудитель черной гнили батата и болезней многих других растений (рис. 84). Большое значение имеют некоторые сапрофиты или слабые паразиты из этого рода, вызывающие синеву древесины (С. pilifera, С. piceae и др.).
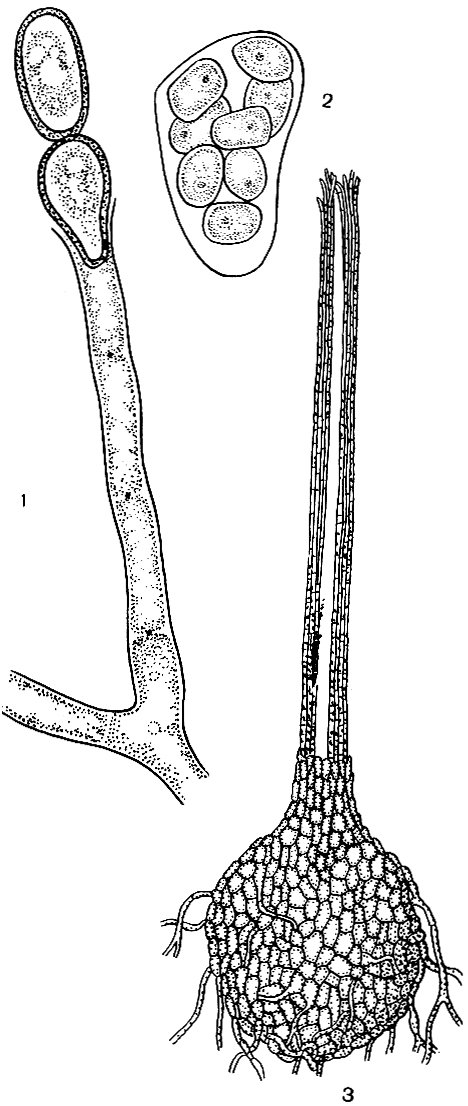
Рис. 84. Цератоцистис бахромчатый (Ceratocystis fimbriata): 1 - конидиальное спороношение; 2 - сумка; 3 - перитеций
Вязовый цератоцистис - один из наиболее известных и важных в хозяйственном отношении представителей рода. Он вызывает так называемую голландскую болезнь вяза, распространенную в Европе и Северной Америке (карта 3) и приносящую большой ущерб лесам, парковым и защитным насаждениям. Болезнь начинается с внезапного пожелтения, увядания листьев и усыхания концов ветвей. При острой форме заболевания дерево усыхает в течение вегетационного сезона, а иногда даже за несколько дней. При хронической болезни листья преждевременно желтеют и частично опадают и пораженные ветви хорошо выделяются на зеленом фоне кроны. Этот характерный симптом голландской болезни получил название "желтого флага". На срезах пораженных ветвей хорошо заметно потемнение сосудов, иногда локализованное, но часто захватывающее всю окружность среза.
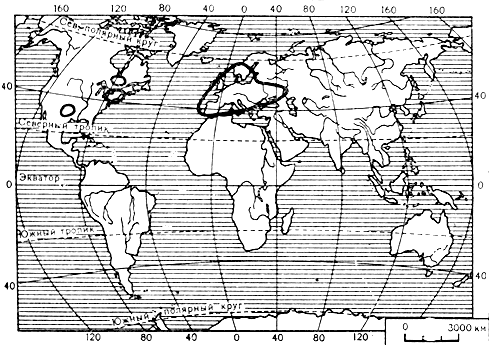
Карта 3. Ареал возбудителя голландской болезни вязов
Голландская болезнь вяза впервые появилась в 1917-1919 гг. в Голландии, а в 1919- 1929 гг. она уже распространилась по Европе, достигнув в 1936 г. западных районов нашей страны. В 1930-1933 гг. она была завезена через порты Атлантики и Мексиканского залива в США и распространилась там в популяциях американского вяза. В 1940 г. она была обнаружена в Канаде. На Американский континент возбудитель был завезен с вязовыми бревнами, пораженными жуками-короедами, переносящими инфекцию.
В Голландии, где в насаждениях преобладает один клон вязов (Ulmus hollandica v. Belgica), некоторые города потеряли до 70% вязовых насаждений. В США потери в насаждениях американского вяза составляли в отдельных районах северо-восточных штатов до 15% в год, а в 1968-1969 гг. в некоторых штатах наблюдалась массовая гибель ильмовых.
К голландской болезни восприимчивы многие ильмовые - некоторые клоны голландского вяза, американский вяз, берест, обыкновенный вяз и др. Хотя полностью иммунные к этой болезни виды вязов не найдены, некоторые из них относительно устойчивы к ней, как, например, перистоветвистый вяз.
В конце апреля - начале мая под отставшей корой больных деревьев в массе развивается конидиальное спороношение гриба - черные коремии типа графиум (Graphium) (рис. 228). Их можно найти также в ходах жуков-короедов и на порубочных остатках - пнях, кучах опилок, торцах бревен, которые служат одним из источников инфекции. Это совпадает с весенним лётом и дополнительным питанием ильмовых заболонников - основных переносчиков возбудителя. Жуки заносят споры гриба в свои ходы, вызывая заражение дерева. Некоторые листогрызущие насекомые (ильмовый листоед, ильмовый слоник и др.) также могут участвовать в распространении инфекции (В. А. Зудилин). Подсохшие споры гриба могут разноситься токами воздуха, распространяться с зараженными древесиной и опилками. Попадая на свежие механические повреждения деревьев, они могут вызвать их заражение. Конидии гриба в древесине сохраняют жизнеспособность более года. Перитеции гриба образуются также в трещинах коры и древесины, в ходах жуков-короедов, но они менее обильны, чем конидиальное спороношение гриба. Интересно отметить, что как конидии, так и аскоспоры гриба погружены в слизь, и это облегчает перенос их насекомыми.
После заражения дерева, уже через 20-24 ч, наблюдается размножение гриба в сосудах ксилемы. Здесь образуется масса микроконидий типа бластоспор, очень мелких, разносящихся по сосудам дерева с восходящими токами. За час они переносятся на расстояние до 15 м, скорость их перемещения определяется интенсивностью сокодвижения. Гифы гриба через поры проникают из сосуда в сосуд. Они распространяются также в сердцевинных лучах и паренхиме дерева. В пораженных клетках образуется темная камедеподобная масса, выделяющаяся в сосуды ксилемы. Причинами увядания дерева считают механическое закупоривание сосудов ветвей и ствола камедью, спорами и гифами гриба, продуктами гидролиза клеточных стенок хозяина и образование грибом токсина.
Борьба с голландской болезнью вяза включает комплекс лесохозяйственных мероприятий (санитарные рубки, обрезка больных ветвей, своевременная уборка порубочных остатков), уничтожение переносчиков болезни, выращивание устойчивых видов и сортов вязов.
Другой важный вид рода - буковый цератоцистис - возбудитель вилта дуба, часто вызывающий массовую гибель деревьев. Болезнь распространена в некоторых штатах США, особенно на Востоке и Среднем Западе. По симптомам она напоминает голландскую болезнь ильмовых. Гриб переносится жуками из рода псевдопидиофторус (Pseudopidyophthorus) при вылете их после перезимовки на пораженных деревьях. У некоторых насекомых он обнаружен внутри тела, в микангиях. Заболевание наблюдается с 40-х годов. До 1952 г. этот вид был известен только в конидиальной стадии - дубовая халара (Chalara guercina).
Перитеции букового цератоцистиса черные, с коротким хоботком, погруженные в сплетение гиф. Половой процесс происходит путем сперматизации, функции спермациев выполняют конидии. Гриб гетероталличен. После дикарионтизации из аскогона развиваются аскогенные клетки, лишенные клеточной стенки. Аскоспоры образуются в результате сегментации протопласта, как спорангиоспоры мукоровых.
В умеренном поясе и особенно в тропиках широко распространен бахромчатый цератоцистис (Ceratocistis fimbriata), вызывающий заболевания многих растений (батата, кофе, гевеи, какао и др.). Он образует типичные для этого рода перитеции, с длинным хоботком, и эндоконидии (рис. 84).
В тропических и субтропических районах земного шара распространены грибы нескольких видов цератоцистиса, вызывающие заболевания важных для этих районов сельскохозяйственных культур. Например, странный цератоцистис (С. paradoxa) широко распространен в тропиках на какао, бананах, ананасах, кокосовой и масличной пальмах, сахарном тростнике, у которых он вызывает гниль.
Интересную группу видов цератоцистиса представляют грибы - возбудители синевы древесины. Цератоцистис пилифера (С. pilifera) вызывает синюю или серовато-синюю окраску заболонной древесины сосны, ели, а иногда и некоторых лиственных - ильма, ясеня и др. Гифы гриба в древесине имеют бурую окраску, поэтому многие считают, что синяя окраска древесины - чисто оптический эффект, основанный на контрасте между светлой древесиной и темными гифами гриба. Однако некоторыми исследователями показано, что в условиях культуры и при росте на древесине этот гриб образует синий пигмент, который вызывает окрашивание древесины. Выделение пигмента грибом происходит в условиях азотного голодания, поэтому окрашивание наблюдается обычно на более поздних стадиях развития гриба в древесине.
Грибы, вызывающие засинение древесины, не могут развиваться при снижении содержания воздуха в древесине до 15-20%.
Обычно грибы синевы не разрушают клеточные стенки и не изменяют механических свойств древесины. Однако считают, что такая засиненная древесина легче поражается дереворазрушающими грибами, хотя эксперименты не всегда подтверждают это положение.
Еловый цератоцистис (С. piceae) часто встречается в Европе и Северной Америке на древесине хвойных, но вызывает лишь слабое ее окрашивание. Меньший цератоцистис (С. minor) вызывает темно-серое или черноватое окрашивание древесины сосны. В ряде стран, например в Швеции,- это один из основных возбудителей синевы древесины. В других странах он не имеет значения как возбудитель окрашивания. Этот гриб встречается на хвойных в ассоциации с короедами, в теле которых содержатся споры гриба.
В США, Японии и ряде стран Европы синеву древесины сосны вызывает цератоцистис ипс (С. ips), встречающийся в ассоциации с короедами из рода Ips.
Образование ассоциаций с насекомыми (древоточцами или короедами) свойственно многим цератоцистисам. У зрелых самок некоторых насекомых имеются даже специальные мешочки - микангии, содержащие споры или дрожжеподобные клетки гриба. Грибы переносятся насекомыми и развиваются в их ходах и прилегающей древесине. Гифы гриба и его споры служат источником пищи для личинок; гриб также размягчает древесину, помогая личинкам проделывать ходы. В свою очередь, насекомые распространяют споры грибов, переносят их на благоприятный для роста субстрат.
Для рода микроаскус (Microascus) из семейства микроасковых (Microascaceae) характерны черные перитеции с коротким носиком, из которых аскоспоры выходят в виде длинных слизистых красно-коричневых шнуров (рис. 85). Йонидиальные стадии обычно типа скопуляриопсис (Scopulariopsis) - аннелоспоры (см. раздел о дейтеромицетах, рис. 230) на разветвленных конидиеносцах.
Микроаскус встречается на разнообразных растительных и животных субстратах, в почве, на экскрементах животных, иногда вызывает заболевания растений (например, М. trigonosporus) и животных (М. manginii, М. cinereus). Нередко один вид обладает очень широкой специализацией к субстратам. Например, треугольный микроаскус (М. trigonosporus) обитает на древесине, в почве, на экскрементах, выделяется из поверхностно стерилизованных семян растений и из очагов поражения при дерматомикозах и онихомикозах. Этот вид образует черные углистые бутылковидные перитеции диаметром 120-250 мкм, с довольно длинным носиком (до 250 мкм) и очень характерные треугольные споры, в массе красно-коричневые.
|
ПОИСК:
|
При использовании материалов проекта активная ссылка обязательна:
http://gribochek.su/ 'Библиотека о грибах'