Отдел Слизевики (Myxomycota) (Т. П. Сизова)
В этом томе, посвященном грибам, описывается небольшая, но интересная группа близких к грибам организмов - слизевиков, или миксомицетов, которая во многих современных системах рассматривается как самостоятельный отдел. Вегетативные тела слизевиков представлены плазменной массой с большим количеством ядер, не одетой оболочкой. Такие тела называют плазмодиями.
Плазмодий - сложное образование. В его составе около 75% воды, а из остальной части около 30% белков; кроме того, в нем содержится гликоген, или животный крахмал, и пульсирующие вакуоли. Некоторые слизевики характеризуются наличием большого количества извести (до 28%) или других включений. У большинства слизевиков в плазмодии находятся пигменты, придающие им самые различные окраски: ярко-желтую, розовую, красную, фиолетовую, почти черную. При этом окраска плазмодия постоянна для данного вида слизевика, но на ее интенсивность очень влияют реакция среды, освещение, температура, питание и другие факторы окружающей среды. Предполагают, что некоторые пигменты представляют собой фоторецепторы, играющие важную роль в развитии слизевиков. Для слизевиков с окрашенными плазмодиями свет необходим для формирования спороношения, которое образуется после периода вегетативного роста.
Размеры плазмодиев у слизевиков разных видов могут быть самые различные, от микроскопически малых (например, Echinostelium minutum, Clastoderma debaryanum) до очень больших. Например, у фулиго (Fuligo) плазмодий может вырасти до нескольких десятков сантиметров в диаметре. Именно такой, большой, ярко-желтый, пенистый, пышный плазмодий этого слизевика изображен в "Жизни растений" К. А. Тимирязева. Вообще плазмодий почти всех слизевиков в течение жизни увеличивается в размерах, и иногда очень быстро, конечно, при наличии благоприятных условий. Так, плазмодий слизевика многоголового (Physarum polycephalum) диаметром 1 см за неделю достиг размера 25 см2!
Основную массу видов слизевиков составляют сапрофитные формы, живущие главным образом в лесах, где-нибудь в глубине гнилых пней, под опавшей корой или опавшими листьями, в трещинах и щелях замшелых колод. До поры до времени плазмодий находится в темноте и питается, главным образом впитывая всей своей поверхностью органические вещества из окружающей его влаги. Но, кроме этого, плазмодий может активно захватывать и твердые пищевые частицы, и живых бактерий, амеб, жгутиковых, мицелий и споры грибов. Собственно, поэтому слизевики нельзя считать чисто сапрофитными организмами.
Плазмодий активно перемещается в направлении источников пищи, т. е. обладает положительным трофотаксисом. Он движется в направлении более влажных мест и навстречу току воды (положительные гидро- и реотаксисы). Пользуясь этой особенностью плазмодия, его можно "выманить", например, из пня. Для этого нужно поместить от края пня в глубь его наклонно полоску стекла, а сверху нее положить фильтровальную бумагу, конец которой погрузить в сосуд с водой. Ток воды может вызвать вползание плазмодия по стеклу, тогда можно не только рассмотреть его под микроскопом, но и проследить, с какой скоростью он перемещается.
Движущие силы токов плазмы в плазмодии еще сравнительно мало изучены. Однако существует предположение, что движение связано с изменением вязкости специального белка - миксомиозина - при взаимодействии с АТФ. АТФ (аденозинтрифосфат) используется во всех реакциях обмена любой клетки живого организма, требующих затраты энергии. Наличие миксомиозина, так же как и АТФ, непосредственно доказано в плазмодии слизевика многоголового. Интересно, что, по-видимому, реакция этих двух веществ протекает так же, как реакция АТФ с актомиозином в мышцах животных и человека.
В прозрачном прикраевом слое цитоплазмы, свободном от органелл, с помощью электронного микроскопа были обнаружены чрезвычайно тонкие нити, находящиеся в непосредственном контакте с оболочкой. Было высказано предположение, что сокращение этих нитей также связано с токами цитоплазмы и движением плазмодия. Токи цитоплазмы в плазмодии можно непосредственно наблюдать под микроскопом. При этом в направлении движения у плазмодия возникают выросты, напоминающие псевдоподии простейших животных, и общий объем цитоплазмы всегда оказывается большим на переднем по движению конце плазмодия. Такая полярность плазмодия, по-видимому, тесно связана с концентрацией калия, т. е. большие концентрации возникают на переднем конце мигрирующего плазмодия. Измерена скорость движения плазмодия. Она довольно значительна, достигая 0,1-0,4 мм в минуту.
Интересно, что при неблагоприятных условиях (большая сухость субстрата, низкие температуры, отсутствие пищи и т. п.) плазмодий может превращаться в утолщенную, твердеющую массу - склероций. Такие склероции могут очень длительно сохранять жизнеспособность и опять превращаться в плазмодий. Известен случай превращения в плазмодий склероция слизевика фулиго, пролежавшего в гербарии 20 лет!
Проследить в природной обстановке цикл развития какого-нибудь слизевика - увлекательное занятие не только для биолога, но для всякого человека, любящего природу. Оказывается, в какой-то момент жизни, определяемый окружающими условиями и главным образом соответствующим состоянием самого плазмодия, отрицательный фототаксис у него меняется на положительный и он сам выползает на поверхность, к свету. Вот тут и можно найти на пнях или просто на земле, на мху слизистые массы различных окрасок - плазмодии. Можно наблюдать за дальнейшим развитием плазмодия на месте или очень бережно, стараясь не повредить, взять его с собой вместе с субстратом, на котором он был найден. Буквально на глазах начнутся чудесные превращения. Весь плазмодий преобразуется в спороношения, различные у разных видов слизевиков. Иногда этот процесс длится всего несколько часов, иногда занимает примерно двое суток.
Интересно, что некоторые группы слизевиков предпочитают какой-то определенный субстрат для формирования спороношения. Так, виды рода бадамия (Badhamia) плодоносят большей частью на коре гнилых деревьев, виды рода крибрария (Cribraria) выбирают древесину хвойных деревьев, виды рода дидимиум (Didymium) предпочитают отмершие листья, а большинство видов рода трихия (Trichia) - мертвую древесину (табл. 1,4).
Самое простое спороношение представляет собой нечто вроде подушечки или лепешки. При его формировании плазмодий, не меняя формы, одевается перепончатой или хрящеватой оболочкой. Более сложные спороношения представляют собой отдельные или скученные плодовые тела, у одних слизевиков - на ножках, а у других - без них (сидячие). У многих эти спороношения очень красивы, особенно при рассматривании в лупу или под бинокуляром.

Таблица 1. Слизевики: 1 - цератиомикса (Ceratiomyxa): общий вид спороношения, справа - часть спороношения со спорами; 2 - ликогала (Lycogala): молодые (розовые) и старые (бурые) эталии; слева - псевдокапиллиций и споры; 3 - арцирия (Arcyria): внешний вид спороношения с капиллицием (в центре), справа - часть капиллиция и споры; 4 - трихия (Trichia): внешний вид спороношения (внизу - часть капиллиция и споры)
У некоторых слизевиков вначале закладываются отдельные спороношения, но на ранних стадиях развития они сливаются вместе и одеваются одной общей оболочкой. Такие образования называются эталиями.
Внутри спороношений развиваются споры. Когда оболочка зрелого спороношения разрывается, споры рассеиваются по воздуху. Таким образом становится понятным выход плазмодия на поверхность перед образованием спороношения. Для образования его необходим свет, а зрелые споры должны распространяться и заселять новые подходящие местообитания.
У многих слизевиков в спороношении имеются особые нити - капиллиций. Они имеют различное строение: ветвистые, неветвистые, в виде полых трубочек, сплошные, похожие на нежную сеть или каркас. Так как на их поверхности большей частью имеются утолщения в виде колец, спиралей, шипиков, бородавок, то они способны к гигроскопическим движениям. У большинства слизевиков они упруго свернуты внутри спороношения, а при вскрытии его выпрямляются, как пружина, меняют положение при изменении влажности воздуха, таким образом разрыхляют споровую массу и содействуют рассеиванию спор.
При благоприятных условиях споры прорастают. Для этого в первую очередь необходима жидкая среда. Если условия неблагоприятны, споры могут очень долгое время, даже несколько десятков лет, сохранять жизнеспособность, не прорастая. На электронных микрографиях спор Didymium nigripes обнаруживаются два слоя (внутренний, более тонкий, целлюлозный, и наружный - хитиновый), а у Physarum gyrosum таких слоев имеется даже три. Видимо, наличие такой оболочки обеспечивает большую сохраняемость жизнеспособности спор.
Если спора прорастает в воде или в питательном растворе, то из пор, имеющихся в оболочке или при разрыве ее, выходят одна, две, иногда даже четыре или восемь зооспор с двумя жгутиками на переднем, суженном конце. Второй жгутик очень короткий, часто согнутый, и появляется он не сразу, поэтому не всегда обнаруживается. В случае прорастания споры просто на влажной поверхности жгутики не вырабатываются и из споры выходят маленькие амебы, так называемые миксамебы. Количество зооспор и миксамеб в среде может увеличиваться, так как они могут размножаться, продольно делясь.
При достижении некоторой критической для данных условий концентрации зооспор или миксамеб наступает половой процесс, т. е. они сливаются попарно задними концами. Их ядра также сливаются, и возникает миксамеба с диплоидным ядром, представляющая собой начало развития нового плазмодия. Ядро его делится без редукции числа хромосом (митотически). Немедленно после митоза в плазмодии начинается синтез ДНК, продолжающийся в течение 1-2 ч. Таким образом в плазмодии увеличивается масса ядерного вещества, ядра синхронно делятся, и плазмодий становится многоядерным, т. е. тем самым диплоидным плазмодием, с которого мы начали рассмотрение жизненного цикла слизевиков.
Следует заметить, что маленькие диплоидные плазмодии могут сливаться друг с другом. Это замечательное явление, ранее толковавшееся как механизм образования плазмодия, имеет весьма существенное значение, так как говорит о вероятном существовании в одном плазмодии ядер различного происхождения, т. е. о возможном гетерокариозе (разнокачественности ядер). Вероятность слияния плазмодиев, имеющих ядра разной генетической структуры, подтверждается тем, что в плазмодии слизевика Didymium nigripes обнаружены ядра с различным количеством ДНК, плоидностью и размерами.
Образовавшийся плазмодий уходит куда-нибудь в глубину пня или под листья, начинает перемещаться, питаться, расти до поры нового спороношения.
В настоящее время многие слизевики удается не только культивировать на искусственных питательных средах, но примерно 40 видов из них осуществляют в этих условиях весь цикл развития. Поэтому слизевики - очень ценны как объекты для различного рода исследований: биохимических, биофизических, физиологических, цитологических и генетических.
Большинство слизевиков - космополиты, т. е. распространены повсеместно, и только некоторые приурочены к тропическим и субтропическим областям, как, например, Physarum nicaraguense и Physarum javanicum. С другой стороны, имеются виды, не встречающиеся за пределами умеренных зон. Таков слизевик Hemitrichia clavata. Немногие виды распространены в альпийских и субальпийских областях (например, Diderma alpinum, Lepidoderma granuliferum, Lamproderma carestiae).
Среди наиболее широко распространенных слизевиков можно назвать вид ликогала - Lycogala epidendrum. Его кораллово-розовый плазмодий образует на мертвой древесине спороношения в виде шариков или горошин от нескольких миллиметров до 1,5 см в диаметре. Вначале они имеют такую же кораллово-розовую окраску, и, если их раздавить, из них вытекает жидкое содержимое такою же цвета (в народе эти спороношения называют "волчье вымя"). Постепенно оболочка спороношения (перидий) буреет, утончается, делается хрупкой и покрывается бородавочками. Наверху образуется отверстие, из которого начинают при малейшем толчке вылетать массы мельчайших спор в виде облачка, заметного простым глазом. Рассеиванию их способствует наличие ветвящегося, уплощенного капиллиция с поперечными складками на поверхности. По внешнему виду такое спороношение очень похоже на обычные грибы-дождевики, внутри которых так же находятся и споры, и капиллиций. Не удивительно поэтому, что, не зная процесса развития этого слизевика, отличного от развития дождевиков, и основываясь на чисто внешнем сходстве, этот слизевик, описанный в XVII в., был отнесен тогда же к грибам, среди которых этот и другие виды слизевиков оставались до середины прошлого столетия. В это время в лаборатории ботаника и миколога А. де Бари был разработан метод онтогенеза, т. е. метод изучения полного цикла развития представителей низших растений. Таким образом была установлена связь между плазмодиальной стадией и спороношением слизевиков. В результате своих работ по слизевикам де Бари пришел к заключению, что они стоят гораздо ближе к амебоидным простейшим животным, чем к грибам. Эту точку зрения разделяют некоторые современные исследователи. Однако большинство микологов считают слизевики не тождественными, но близкородственными к грибам, среди которых также имеются представители с вегетативными телами, выраженными как голая плазменная масса.
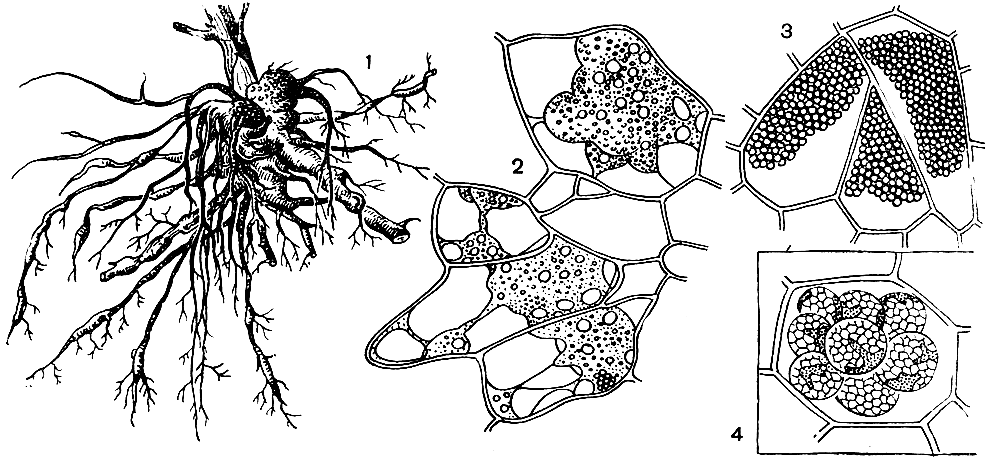
Рис. 1. Кила капусты и порошистая парша картофеля: 1 - общий вид пораженных килой корней; 2 - клетки корня с плазмодием возбудителя килы; 3 - клетки корня со спорами паразита; 4 - губчатые комочки спор возбудителя порошистой парши картофеля в тканях клубня
К вопросу о происхождении слизевиков мы еще вернемся, а сейчас приведем очень краткие описания некоторых слизевиков из наиболее часто встречающихся, как, например, уже упоминавшиеся фулиго и ликогала.
Род слизевиков с экзогенно (наружно) образующимися спорами (односпоровыми спорангиями) - цератиомикса (Ceratiomyxa), а один из широко распространенных видов этого рода - Ceratiomyxa fruticulosa. Плазмодии его прозрачные, бесцветные или желтоватые, а возникающие из них спороношения снежно-белые, очень нежные; они представляют собой пучки колонок или сосочков высотой 1-2 мм, поверхность которых покрыта спорами, сидящими на тонких ножках (табл. 1,1).
По количеству известных видов (более 80) первое место среди слизевиков занимает род физарум (Physarum, табл. 2,5).

Таблица 2. Слизевики: 1 - дидерма (Diderma): внешний вид спороношения, внизу - часть капиллиция и споры; 2 - стемонитис (Stemonitis): внешний вид спороношения, слева - часть сетчатого капиллиция и споры; 3 - физарум (Physarum): общий вид спороношения, внизу - часть капиллиция и споры; 4 - леокарпус (Leocarpus): общий вид спороношения, внизу - часть капиллиция и споры
Плазмодии представителей рода физарум бесцветные или окрашены в желтые либо оранжевые тона. Спороношения очень разнообразны по окраске и строению, сидячие или на ножках, открывающиеся неправильными разрывами оболочки. Для них характерно наличие извести в оболочке и в узлах капиллиция.
На древесине часто встречаются виды рода стемонитис (Stemonitis). Их плазмодий, белый или лимонно-желтый, преобразуется в спороношение в виде тесно скученных изящных "перышек", представляющих собой спорангии удлиненной формы, которые имеют ножку, продолжающуюся в колонку внутри спорангия. Тонкая сеть капиллиция соединена с колонкой. Перидий (оболочка спорангия) исчезает очень рано, и остается сеть капиллиция, коричневая от покрывающих ее спор (табл. 2,2).
Спороношения видов рода арцирия (Arcyria) также красивы и изящны. Они отличаются более яркими окрасками (например, карминово-красной, желто-бурой) сети капиллиция, выходящей из "чашечки" - остатка перидия в верхней части ножки (табл. 1,3).
Очень эффектны спороношения Leocarpus fragilis. Они похожи на крошечные сливы с темной, блестящей, как бы лакированной оболочкой (табл. 2,4).
Все указанные здесь примеры не исчерпывают разнообразия в строении слизевиков и могут дать только некоторое представление об отдельных родах, встречающихся повсеместно.
Кроме вышеописанных, имеется еще одна очень важная группа слизевиков - внутриклеточные паразиты растений. Вегетативное тело их представлено также плазменной массой с большим количеством ядер, преобразующейся затем в споры. В отличие от свободноживущих сапрофитных слизевиков они не образуют никаких специальных спороношений, так как вместилищем для спор служит оболочка клетки растения-хозяина.
Важнейший представитель этой группы - возбудитель килы капусты и других крестоцветных - Plasmodiophora brassicae. Корни больного растения имеют необычный, уродливый вид (рис. 1).
Они вздуты, и их общая масса составляет около половины общей массы растений вместо 1/30 или 1/70 у здоровых растений. Если разрезать корни, находящиеся на ранней стадии заражения, то в некоторых клетках их видны миксамебы или плазмодии слизевика. На поздней стадии заражения эти клетки корня оказываются буквально набитыми мелкими круглыми спорами, на которые распался плазмодий.
Интересно, что клетки корня, содержащие паразитов, сильно увеличиваются в размерах по сравнению с нормальными и, кроме того, усиленно делятся, в результате чего образуется опухоль. Если, например, заражается капустная рассада, кочаны могут совсем не образоваться, а при заражении взрослых растений капусты кочаны недоразвиваются. Корни, сгнивая, освобождают массу спор, попадающих в почву. Распространению спор в почве способствуют дождевые черви, различные почвенные насекомые, токи воды и т. п. Человек также может содействовать их расселению и переносу на новые места, например, применяя в качестве удобрения навоз животных, которым скармливался зараженный турнепс. В почве споры могут долго сохраняться, прорастая при благоприятных условиях; выходящие из них двужгутиковые зооспоры или образующиеся из зооспор миксамебы заражают новые растения, главным образом через корневые волоски. Внутри клеток корня, при слиянии миксамеб и последующих митотических делениях ядра, образуются многоядерные плазмодии.
Это заболевание капусты было известно издавна и наносило большой ущерб овощеводству. Так, в иные годы погибало более половины урожая капусты. Однако причину болезни искали то в повреждениях насекомыми, то в неблагоприятных условиях почвы и т. п.
В 1872 г. Санкт-Петербургское общество садоводства объявило премию за выяснение причин болезни и разработку мер борьбы с нею. Независимо от объявленного конкурса изучением загадочного заболевания капусты занялся М. С. Воронин, который описал уже в 1875 г. в главных чертах цикл развития возбудителя капустной килы и предложил меры борьбы с паразитом (прожигание почвы в парниках, отбор рассады, плодосмен). Эта работа М. С. Воронина вошла в фонд классических исследований по болезням растений.
Многие детали были впоследствии уточнены работами русских и зарубежных исследователей. В частности, были выяснены условия, особенно благоприятствующие прорастанию спор и заражению растений: высокая влажность почвы (в пределах 80-90% полной влагоемкости), температура почвы 18-24 °С, рН 6,0-6,5.
Особенно вредоносна кила капусты в европейской части СССР (на подзолистых почвах нечерноземной зоны). Однако в последние годы болезнь обнаружена в ряде восточных районов (Сибирь, Дальний Восток) и в некоторых южных районах (Азербайджан и др.), особенно при выращивании капусты в поймах рек.
Оказалось, что Plasmodiophora Brassicae может заражать примерно 200 видов культурных и дикорастущих крестоцветных. Возможно, последние могут служить местом резервации паразита.
Другой представитель этой группы, также имеющий практическое значение,- возбудитель порошистой парши картофеля - спонгоспора - Spongospora solani, поражающая клубни, корни, реже столоны картофеля, томаты и другие виды пасленовых. Многоядерные плазмодии, локализующиеся в покровных клетках клубня картофеля, образуют там губчатые комочки плотно спаянных спор. Пораженные клетки отделяются от нижележащей ткани клубня слоем пробки. Вначале пораженный участок имеет вид бугорка на клубне. Потом на этом месте кожица клубня звездчато разрывается и образуется язвочка с коричневым порошистым содержимым из скопления губчатых комочков спор (рис. 1). Корневая форма имеет вид небольших белых желвачков, потом темнеющих и распадающихся. В почве из спор выходят зооспоры, проникающие в корневые волоски или непосредственно в клетки эпидермиса. Если язвочки находятся вблизи ростков, то последние могут не развиваться. Кроме того, в эти язвочки могут попадать микробы, разрушающие клубень.
Заболевание известно почти во всех странах умеренного пояса. В СССР оно распространено в Белорусской ССР и Ленинградской области. Главные меры борьбы - соблюдение севооборотов и отбор здоровых клубней для посадки.
Очень интересна среди низших организмов группа акразиевых (Acrasieae), развивающихся главным образом на навозе. На определенной стадии развития акразиевые внешне очень похожи на плазмодии и, как и слизевики, образуют спороношения. Однако при знакомстве с циклом развития их становится ясно, что это сходство чисто внешнее. Во-первых, у них совершенно отсутствует подвижная жгутиковая стадия - зооспоры. Во-вторых, плазмодий у них не настоящий. В вегетативной фазе развития акразиевые похожи на амеб, многие передвигаются при помощи псевдоподий, размножаются делением, и количество их таким образом увеличивается. Когда исчерпываются запасы пищи, амебы сползают вместе и сливаются в общую массу, не теряя при этом своей индивидуальности, т. е. слияние это не полное. Эта единая масса амеб, внешне очень напоминающая плазмодий, называется псевдоплазмодием.
Сходство акразиевых со слизевиками увеличивается и в том, что псевдоплазмодии некоторых акразиевых начинают активно двигаться (мигрировать) и затем образуют спороношения. У наиболее известного представителя акразиевых - Dictyostelium discoideum, встречающегося на навозе,- спороношение белого цвета, ножка прямая, длиной 3-8 мм, несущая на вершине шаровидную головку спор диаметром 0,2-0,3 мм. При формировании спороношения одни амебы превращаются в клетки ножки, а другие, вползая по ним кверху, образуют споры. И те и другие вырабатывают при этом целлюлозные оболочки (рис. 2).
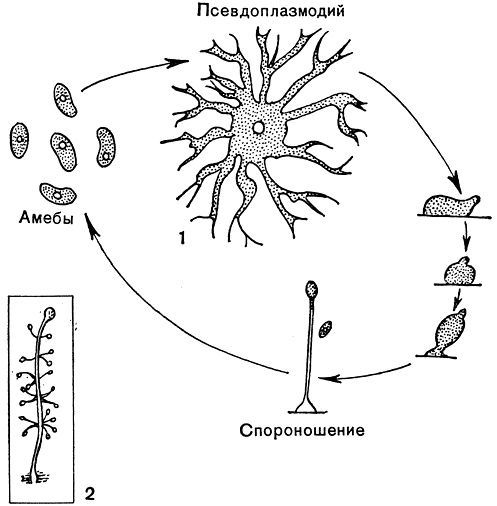
Рис. 2. Акразиевые: 1 - Dictyostelium discoideum: фазы развития; 2 - Polysphondilium violaceum: спороношение
У Polysphondilium violaceum спороношение фиолетового цвета, длинная ножка мутовчато ветвится и каждая веточка кончается споровой головкой удлиненной формы (рис. 2). При прорастании из спор снова выходят амебы.
Многие акразиевые хорошо развиваются в культуре. Например, Dictyostelium discoideum проходит весь цикл развития за 3-4 дня, если его посеять на агаризированный отвар сена с бактерией кишечной палочкой (Escherichia coli). Это делает их очень удобными объектами для различного рода исследований.
Мы рассмотрели три группы внешне похожих организмов. Но имеются ли у них родственные связи или общие предки? Сапрофитные слизевики правильнее всего считать самостоятельной ветвью развития, возникшей из простейших жгутиковых и рано обособившейся в процессе эволюции. Их плазмодий можно рассматривать как результат приспособления к наземной жизни, так как у многих современных бесцветных и окрашенных жгутиковых в определенных условиях, например при подсыхании водоемов, в которых они живут, также возникает плазмодий. Появление спороношения в цикле развития представляет дальнейший шаг эволюции в сухопутных условиях.
К сапрофитным слизевикам тесно примыкают паразитные слизевики, отличающиеся главным образом отсутствием морфологически обособленных спороношений, что можно объяснить их образом жизни в качестве внутриклетных паразитов.
Что касается акразиевых, то вероятнее всего предположить их родство со свободноживущими амебами. Однако своеобразие этой группы представляет еще очень много загадочного.
|
ПОИСК:
|
При использовании материалов проекта активная ссылка обязательна:
http://gribochek.su/ 'Библиотека о грибах'